
Jellyfish
セクションアウトライン
-
-
We have been studying the muscle proteins of invertebrates (such as the troponin-tropomyosin system). However, we started biochemical research on jellyfish proteins six years ago when a student who was interested in jellyfish came to study with us.
The jellyfish is a very attractive research subject:
● Many things about jellyfish remain unknown, although it looks like a simple animal.
● Depending on the species, jellyfish can be cloned and its breeding and maintenance are easy.
If you are interested in protein research or like jellyfish, would you like to study in our laboratory?
The description of the course is slightly biased, but please contact us at tanaka-h@fish.hokudai.ac.jp if you have any comments, suggestions, or questions.
-
-


Jellyfish are classified as primitive invertebrate cnidarians. Can they really be considered primitive? Even though they may be primitive, their bodies have a variety of functions. By learning about the mechanisms of jellyfish bodies, we can get a glimpse into the fascinating world of biological evolution. I hope that making as many people as possible aware of biological evolution and the fascination of biodiversity will help us achieve the goal 14 LIFE BELOW WATER.
The United Nations has designated the decade starting in 2021 as the "Decade of Ocean Science," with the aim of contributing to the SDGs.
-
-
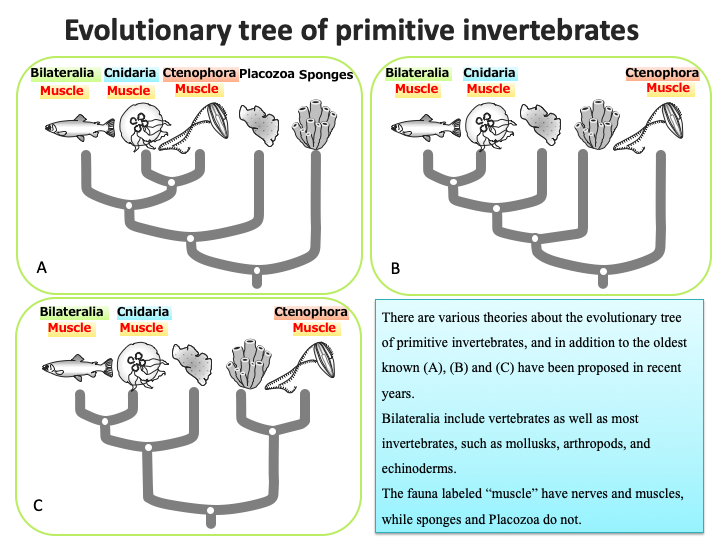
Cnidaria and Bilateralia have much in common regarding the mechanisms of action of the muscles and nerves at the molecular level. In contrast, the Ctenophora appears to be very different from these, and its neurotransmitters are different from those of other animals.
Some jellyfish belong to the phylum Cnidaria (including coral, sea anemone, and hydra), such as Aurelia aurita, Mastigias papua, and Aequorea victoria, while others belong to the phylum Ctenophora, such as Beroe cucumis. Historically, these were grouped together as the phylum Coelenterate, but genetic analyses have recently revealed that they belong to very different groups of animals (B and C in the figure below).
In addition, jellyfish are thought to be the first animals to have acquired nerves and muscles approximately 600 million years ago. Animals that have acquired nerves and muscles are able to “act” even when they grow in size, adapting to various environments, dramatically evolving, and developing intelligence. Jellyfish muscle is composed of the actin-myosin system that, albeit primitive, efficiently uses intracellular energy (ATP) to generate large forces. In addition, its nerves have the division of roles corresponding to the sensory nerves that receive and transmit stimuli such as light and contact, motor nerves that control muscle contractions, and autonomic nerves that regulate the functions of various organs.
Jellyfish have a scattered nervous system and do not have a brain. However, on observation, they sometimes behave as if they were thinking and acting (Video: Predatory behavior of Eutonina indicans and polyp). The rhythm of the beat of Cnidarian jellyfish is created by the nerve cells located on the edge of the umbrella. In addition, the edge of the umbrella contains a sensory organ that senses light and gravity, and some species have eyes with lenses and retinas. Therefore, nerve cells are concentrated in a ring shape on the edge of the umbrella, which is thought to work like the central nervous system.
-
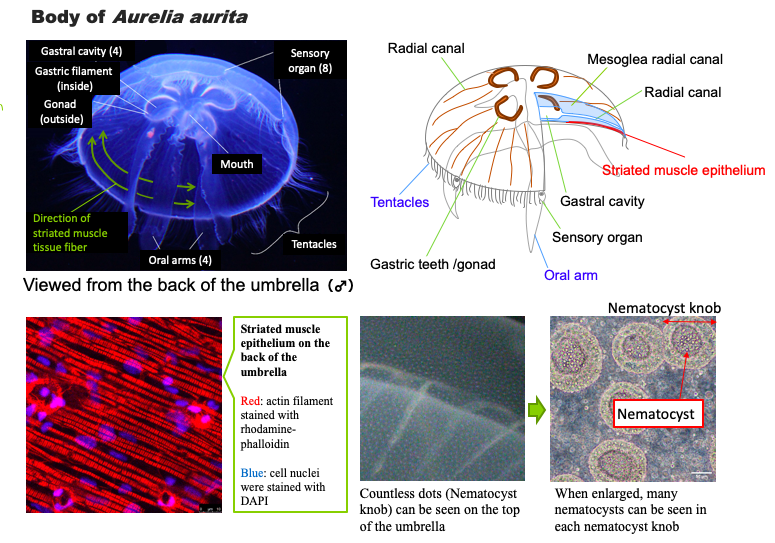
The digested and liquefied food is carried throughout the body via radial canals that radiate from the gastral cavity. The radial canals not only act as a nutrient absorbing organ like the intestinal tract, but also circulate nutrients throughout the body like blood vessels. However, they are not the equivalent of the heart, and the flow of fluid in the radial canals is caused by the movement of the cilia that grow inside. While the flow direction in some radial canals is from the gastral cavity to the edge of the umbrella, others have the flow from the edge of the umbrella back to the gastral cavity. The fluid that returns to the gastral cavity is thought to contain waste products and undigested and unabsorbed food, but these (excrement) are released again into the sea through the oral arms. In the oral arms, the route through which food passes is different from the route through which excrement passes.
The majority of the body of Aurelia aurita is comprised of collagen gel (mesoglia) that is rich in water (95%). Amoeboid cells are sparsely present in the mesoglia, which appear to be responsible for biological defense, gel repair, and nutrient transport. The surface of the mesoglia is covered with one or two thin layers of cells, in which major vital functions, such as the nerves and muscles, digestion and absorption, and germ cell formation, are concentrated.
There are tentacles on the edge of the umbrella to catch food, and the surface of the tentacles is lined with “nematocysts,” which are capsules containing poisonous stings. When zooplankton, a prey, comes into contact with the nematocysts, a poisonous sting is ejected vigorously, piercing the body of the prey to inject the venom. The killed prey is entangled in the mucus secreted from the body surface and collected on the edge of the umbrella by the movement of the cilia on the body surface. The collected food is further licked by the oral arms and carried through the oral arms to the mouth located in the center of the underside of the umbrella. Jellyfish often have four gastral cavities immediately above the mouth, and because there are gastric filaments and gonads along the outer edge of the gastral cavity, a four-leaf clover-like shape is seen in the center of the transparent body. The gastric filaments look like short, dense hair bundles, and the trapped food is digested by secreted digestive enzymes. Although individuals with six gastral cavities are common, some have three, five, or seven, in which there are six, three, five, and seven oral arms, respectively.
-
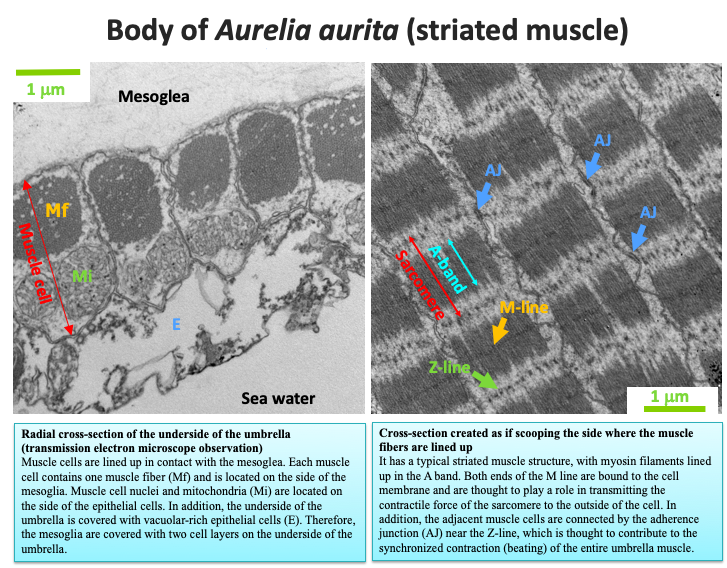
The beating of a jellyfish umbrella is caused by the contraction and relaxation of the muscles in the epithelium behind the umbrella. There are fibers of striated muscle on the back of Aurelia aurita umbrella that run concentrically. When they contract, the umbrella becomes deflated, and when they relax, the elasticity of the thick mesoglia above the umbrella restores them to the original open shape. Jellyfish can swim in the water by repeating this action. Aurelia aurita at the ephyra larval stage has radial muscles, which run radially, in addition to the concentric circular muscles, but the radial muscles degenerate as they grow. On the other hand, the radial muscles remain even after maturity in Cyanea capillata. The beating cycle becomes longer (slower) as it grows, and it changes depending on the water temperature. With individual differences in the beating cycle, those jellyfish beating the umbrella vigorously grow well because their tentacles on the edge of the umbrella flutter well with each beat, collecting and eating a lot of food.
Each muscle fiber is contained in a muscle cell. Muscle fibers contain actin filaments and myosin filaments, and myosin, the main protein component of the myosin filaments, binds to the actin filaments, pulling them, as it decomposes ATP. Then, the actin and myosin filaments slide against each other, causing the muscle fibers to contract. The striated muscle of A. aurita has a striated structure similar to the striated muscle of the Bilateralia, despite some different characteristics. One of the differences is that the length of its sarcomere, which is a repeating unit of striated muscle, is shorter (1.5 micrometers) than those of many Bilateralia (2.5 micrometers). Comparing the case in which muscle fibers of the same length are composed, there are those with many short sarcomeres, while there are a small number of long sarcomeres, and the shorter sarcomere muscle fibers have a higher overall shortening rate even if the actin and myosin filaments slide at the same rate. In other words, the striated muscle of Aurelia aurita is thought to have a mechanism that allows the entire muscle to contract quickly, even if the actin and myosin filaments slide at a low speed. Another characteristic is the Z‐line morphology, which looks very dark in the Bilateralia when viewed under an electron microscope. However, it gives the impression of discontinuity and fragility in jellyfish.
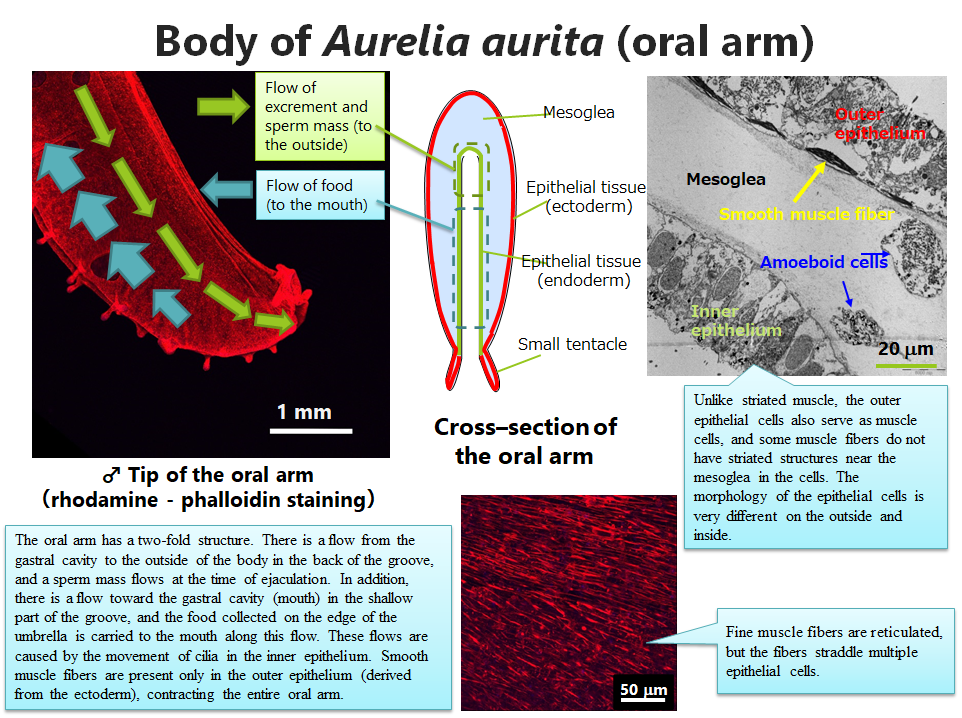
On the other hand, the tentacles and oral arms have smooth muscles. The oral arms are folded in half, and food, excrement, eggs, sperm, and inedible objects caught by mistake move on the inner side. These take different routes: sperm are sent from the testes to the tip of the oral arm through the innermost part of the double-folded groove and released into the sea, while the eggs exit the ditch near the mouth. These are caused by the movement of the cilia in the inner epithelium. The inner and outer epithelia are quite different in their cell morphology, and smooth muscle fibers are located in the outer epithelial cells. This is very different from the striated muscle, which has separate epithelial cells and muscle cells. Smooth muscle fibers appear to be reticulated and connected together throughout the oral arm, but are actually divided into individual epithelial cells.
-
The “nematocysts” that characterize the Cnidaria are the capsules containing poisonous stings with a highly delicate mechanical structure. Nematocysts are used not only to catch prey and protect the jellyfish from foreign enemies, but also to adhere to the base. Jellyfish belonging to the Ctenophora do not have nematocysts, but some instead have “colloblasts” that capture prey by adhesion. In addition, some Ctenophora and mollusks are known to place the nematocysts of captured Cnidaria on their body surface for their protection. The presence of nematocysts is the cause of “being stung by jellyfish” in humans and has attracted great attention. However, the molecular mechanism by which the poisonous stings are ejected from the nematocysts is still poorly understood.
The nematocyst itself is not a living cell but a structure that is created inside the cnidocyte and pushed out of the cell by exocytosis. In Aurelia aurita, nematocysts are present not only throughout the body surface but also inside the gastral cavity. In addition, nematocysts are most abundant on the tentacles, but they are also abundant on the upper surface of the oral arm and umbrella. Surprisingly, there are relatively few nematocysts on the back of the umbrella.



