
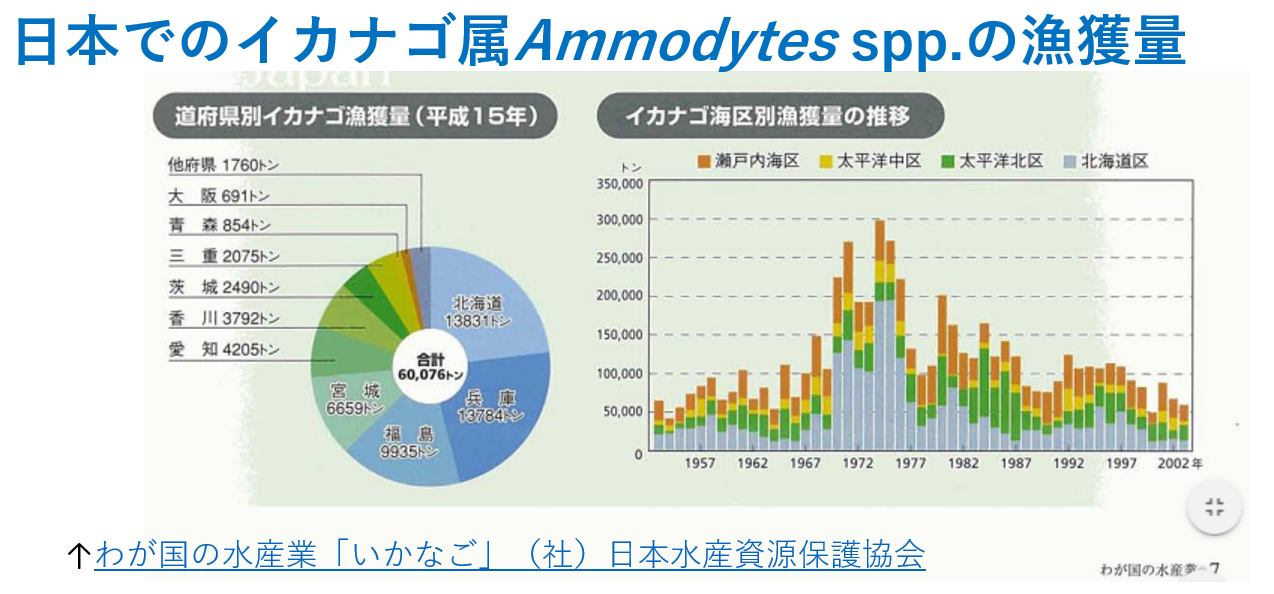
陸奥湾のイカナゴ属(Ammodytes sp.)の生活史
Section outline
-
-



海を豊かに保ちつつ、漁業をすることが大事です。私たちが漁獲してもよい水産資源量はどのように決まるのでしょうか。人間が漁獲すれば水産資源は減少するでしょうが、自然現象でも水産資源の量は大きく変動します。とくに、仔魚期における生残率に着目する必要があります。これらを正確に理解することを通して、SDGs(産業と技術革新の基盤をつくろう、海の豊かさを守ろう)に貢献したいと考えています。
国連では2021年からの10年間を、「海洋科学の10年」に定めて、SDGsに貢献することを目指しています。国連が定める海洋科学には、水産漁業の分野も含まれます。
-
-
-
イカナゴ属は日本列島周辺に3種生息します。

1. イカナゴ Ammodytes japonicus
瀬戸内海・伊勢湾・若狭湾・陸奥湾・三陸沿岸・北海道周辺に生息
2. オオイカナゴ Ammodytes heian
陸奥湾・三陸沿岸・北海道周辺に生息
3. キタイカナゴ Ammodytes hexapterus
北海道周辺のうち,石狩湾・稚内周辺に生息(陸奥湾には生息しない)
1と2は,2015年以前は「イカナゴ Ammodytes personatus」と表記されていましたが,DNA解析の結果,2種が混じっていたことがわかりました。DNA解析でのみ区別できます
キタイカナゴと1-2の区別は,産卵期や背鰭軟条数でおおよそは区別できますが,実際には外見からの区別はほとんど無理です。本種もDNA解析が望ましいです。
2015年以降,A. personatusはベーリング海からカリフォルニア沖の太平洋沿岸に生息し,日本周辺には生息しないことになりました(Orr et al., 2015)。
シワイカナゴ Hypoptychus dybowskii という紛らわしい名前の魚がいますが,この種はトゲウオ亜目Gasterosteoideiの仲間です。
-
-
一般的に、
- 卵やから孵化した直後を仔魚と呼びます。イカナゴ属の1種も含めて,多くの海産魚類は親魚とは似ていない脆弱な形態です。全身の骨格は軟骨です。
- その後成長するにつれて,脊索骨は頭部から尾部に向かって硬骨に変化してゆきます(化骨ossificationと言います)。
- 逆に鰭は尾部から頭部に向かって鰭条が完成してゆき,最後に胸鰭が完成する魚が多いです。全部の鰭の鰭条数が親魚と同じになると,変態が完了し,名前が稚魚に変わります。
- 多くの仔魚は,プランクトン(浮遊)生活者です。
イカナゴ属の小型仔魚、大型仔魚、稚魚の区別です。



-
-
-
漁船は夜に灯りをつけて,浮上してくる稚魚を網で掬って漁獲します。この稚魚は「小女子(こうなご)」とも呼ばれ,天日で干して出荷されたものが「こうなごのちりめん」で,関西では甘い「釘煮(下写真)」にします。

-
-
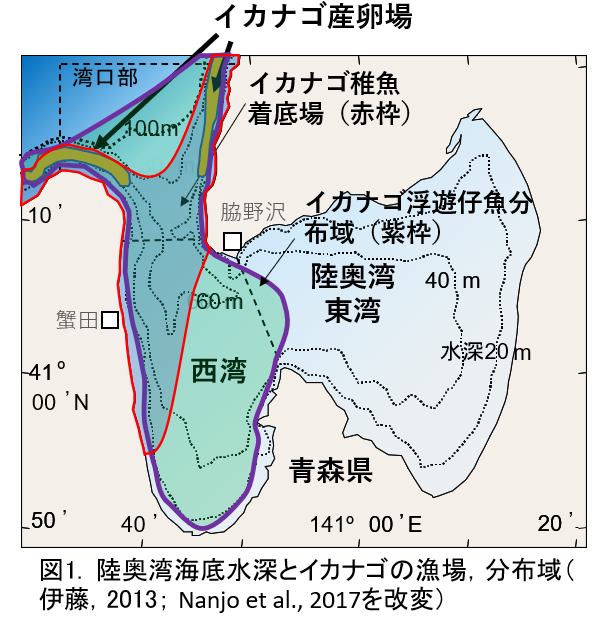
- 青森県の陸奥湾には,湾口部海底にイカナゴ属の1種(Ammodytes sp.)の産卵場があり,2月頃に孵化した仔魚は浮上して陸奥湾西湾でプランクトン生活を送ります。
- その後5月に稚魚まで成長すると,昼間は西湾西側から湾口部の着底場で,海底上に暮らすようになります(図1)。
-
イカナゴ属の1種は2~5歳で成熟し,毎年雌1尾当たり2.2~6.6万個産卵します。これらの卵のうち,雄1尾と雌1尾の合計2尾が漁獲をかいくぐり,自然死亡もなく,成魚になって産卵に参加すれば,イカナゴ資源は維持できるはずです。
産卵数から計算するとその生残率は0.0031~0.0089%であり,宝くじに大当たりするくらい低い確率です。
小型で脆弱な卵や仔魚・稚魚の時期は,未成魚期や成魚期に比べて生残率が低いです。これを「初期減耗」といいます。また,わずかな環境要因の変化が,初期減耗に大きな影響を及ぼします。初期減耗が生じる原因には様々な説があります。例えば,
飢餓仮説:小型仔魚は遊泳速度が低いために,餌密度が低いとお腹を空かせて死亡することがあります。
輸送仮説:小型仔魚は遊泳能力が乏しいプランクトン生活者なので,ネクトンのように生活しやすい水温層や生育場に泳ぎ着くことができません。異常海流で生残できないような外洋に流されてしまうと,生残率が下がります。
水温仮説:急激な水温低下や高水温化が生じると,逃げ切れずに大量死が起こります。
-
-
-

方法1:北大水産学部附属練習船うしお丸と,地元の漁船をチャーターして,プランクトンネットでイカナゴ属の1種の仔魚と,その餌生物であるプランクトンを採集。
方法2: CTDという海洋観測機器で,水温・塩分,海水密度を2週間おきに3年間調査。
-
-
-
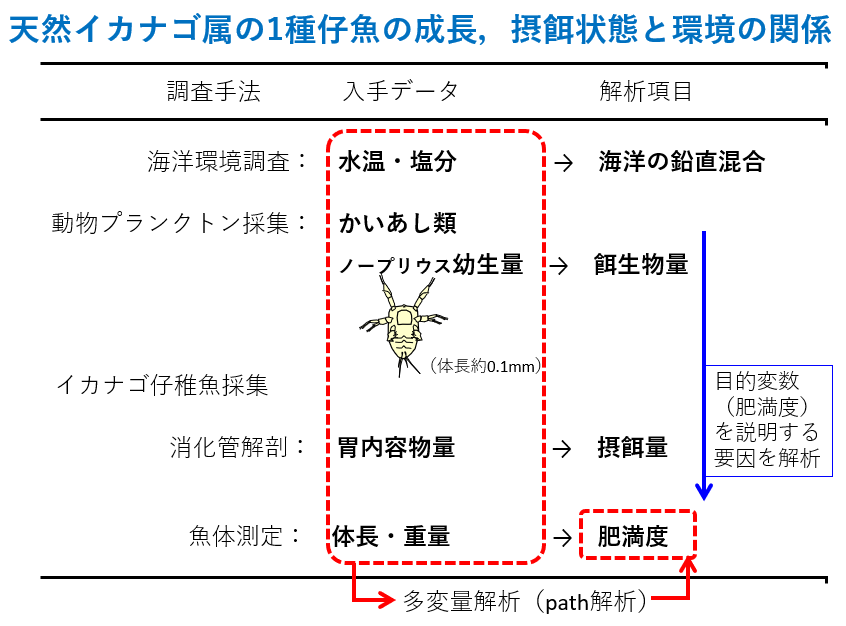
天然イカナゴ属の仔魚の成長、摂餌状態と環境の関係を調べるための、調査手法、入手データ、解析項目です。
-
-
Path(パス)解析の結果(図3)
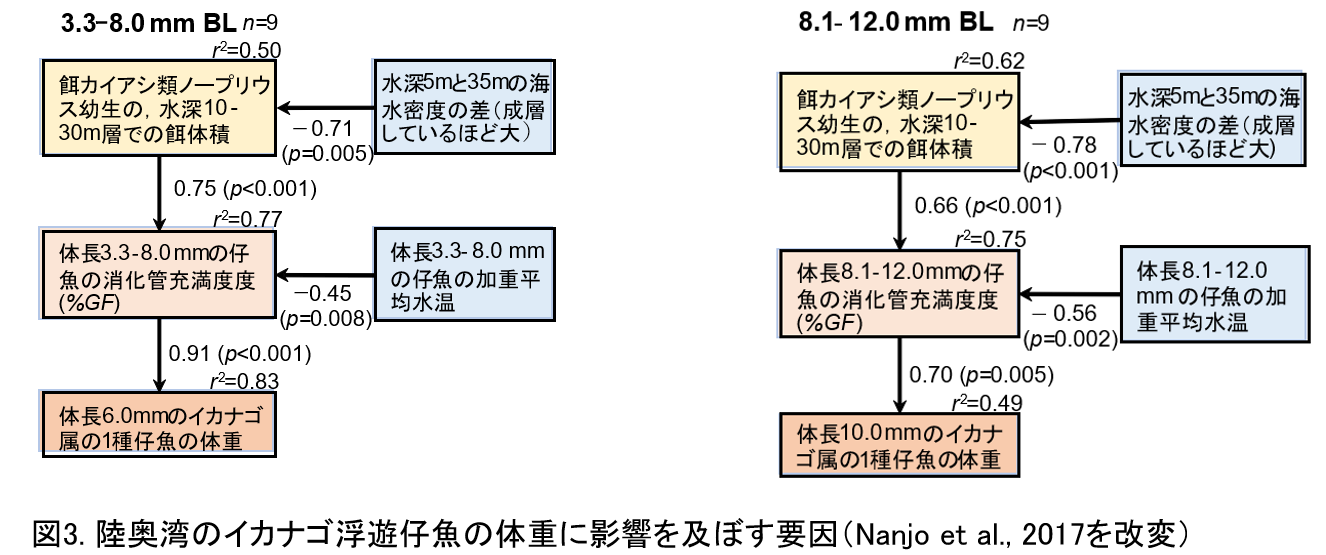
結果1:念のため小型仔魚(上段)と大型仔魚(下段)に分けて解析しました。矢印の横や下の数字は,パス係数β(方向のある相関係数のようなもの)で,正の値で正の相関,負の値で負の相関があると考えてください。
-
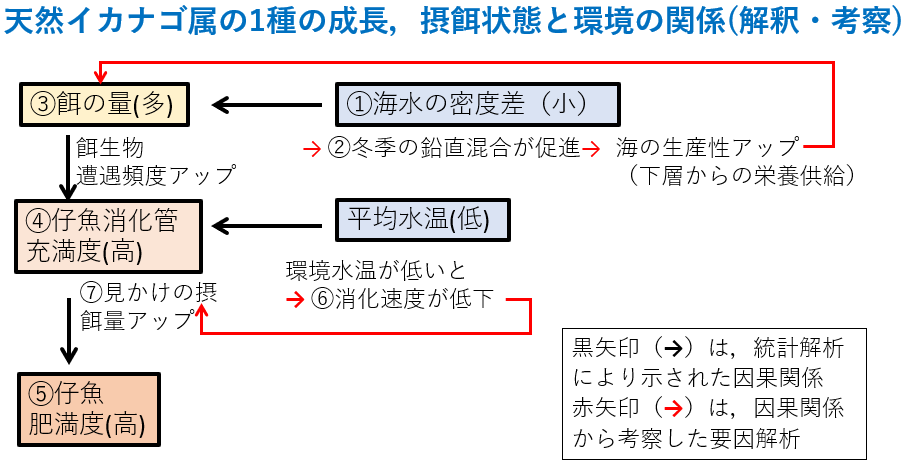
①水深5mと35mの海水密度の差が小さいと(つまり陸奥湾の表層に陸水流入量が少ないと),
②冬季の鉛直混合が促進されて,
③餌生物が増え,
④餌を多く食べ,
⑤体長6mmと10mm時点で栄養を蓄えて太っている
これらが統計的に証明出来ました。
ただし低水温だと、
⑥消化速度が低下して,
⑦見かけ上摂餌量がアップしてエネルギー取り込み量はむしろ低下しますが,
③の効果(β=0.75と0.66の絶対値)>⑥の効果(β=−0.45と−0.56の絶対値)なので,
陸奥湾の水温環境は⑤肥満度を低下させるほど強い効果ではないことが統計的に証明出来ました(0.60倍(=0.45÷0.75)から0.85倍(=0.56÷0.66)の効果と計算できます)。
-
陸奥湾のイカナゴの生残率の年ごとの違いを示します。
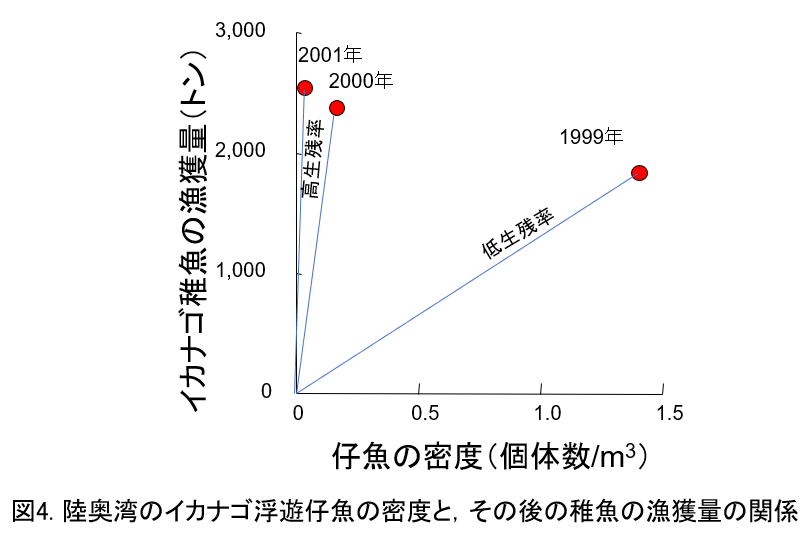
- 1999年は3年間で一番仔魚が高密度だったのに,稚魚まで生残した個体は一番少なかった。つまり,仔魚から稚魚に至るまでに著しく減耗したことを意味します。図中の直線の傾きが小さいので,1999年は低生残率だったといえます。
- 2001年は逆に高生残率だったといえます。
-
-
降水量の年変化を示します。
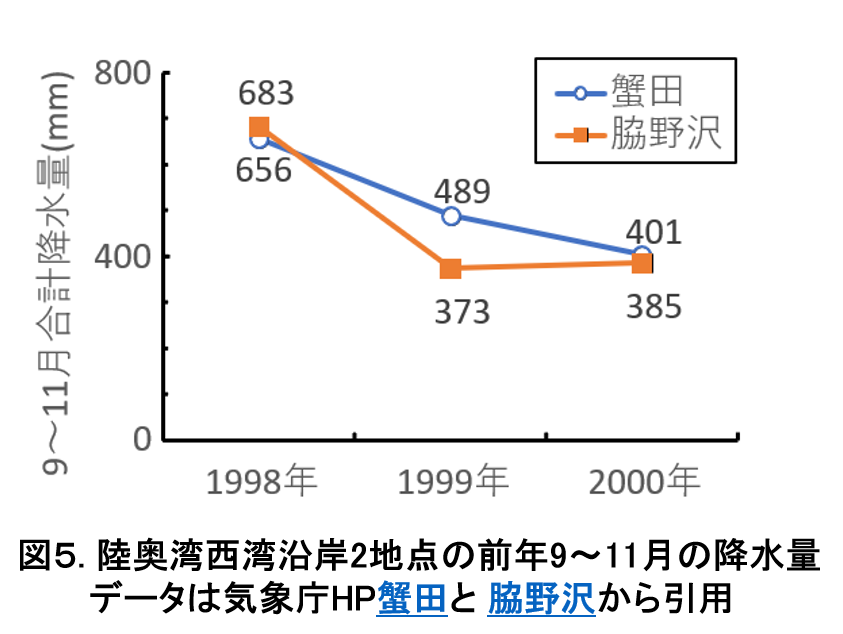
1999~2001年の3のうち1999年は,前年秋季(1998年9~11月)に大雨による降水が多量に陸奥湾に流入していました(図5)。
環境要因とイカナゴ栄養状態の関係(解釈・考察)前年秋期に陸奥湾の表層に陸水流入量が少ないと,冬季の鉛直混合が促進されて餌生物が増え,餌も多く食べることができ,体長6 mmと10 mm時点で栄養を蓄えて太る。
逆に,陸水流入が多いと,イカナゴが痩せる(栄養状態が悪い)。
-
-
-
考察:結果1~3を総合して,
「秋に雨が多い年には,春にイカナゴが減る」と予測できるようになりました。(理論的な「風が吹けば桶屋が儲かる」です)
こういった年には,獲り過ぎないように漁獲量を抑制する必要があるといえるでしょう。
図4で示された仔魚の密度はどのように決まるのでしょうか?卵の量は基本的に雌親の総重量に比例しますので,雌親魚の栄養状態が毎年同じなら,親の量で卵の量は決まります。
しかし仔魚の生残率はここで調べたように自然現象によって大きく変動し,人間がコントロールできる余地はありません。
せめて人間ができることは(種苗放流を除けば),獲り過ぎないようにして雌親魚を増やすことだけです。また禁漁すれば雌親魚が高齢・大型化するので,産卵数も増えます。そして仔魚の生残率が高くなる年を待つしかありません。



