
数値流体力学的を用いた平行移動する魚の流体力学的効果
Section outline
-
-

表1. 試験魚のパラメータ
-

図1. 体軸上の点
-
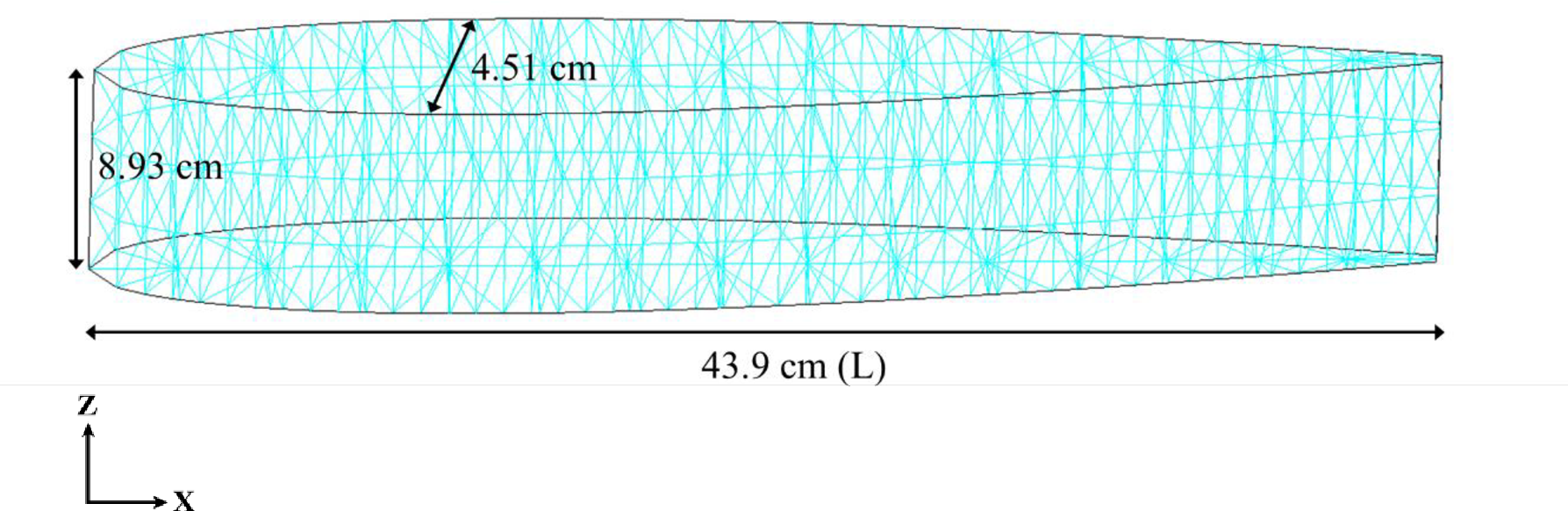
図2. CFD解析に使用したモデル
-
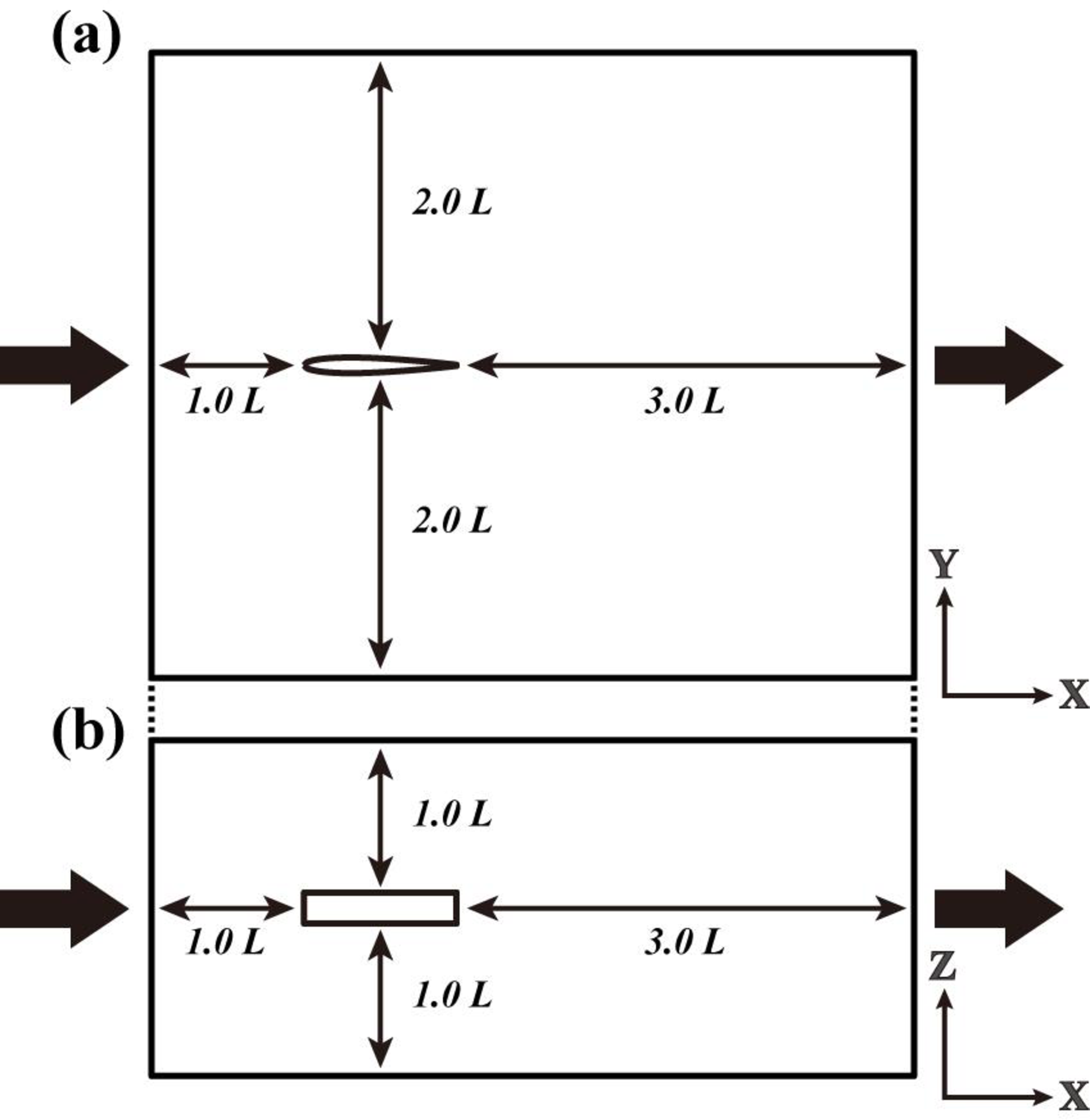
図3. ソリタリースイミングの CFD 解析のための解析領域。(a) 上面図、(b) 側面図。
-

図4. 孤立遊泳の CFD 解析におけるメッシュ分割
-
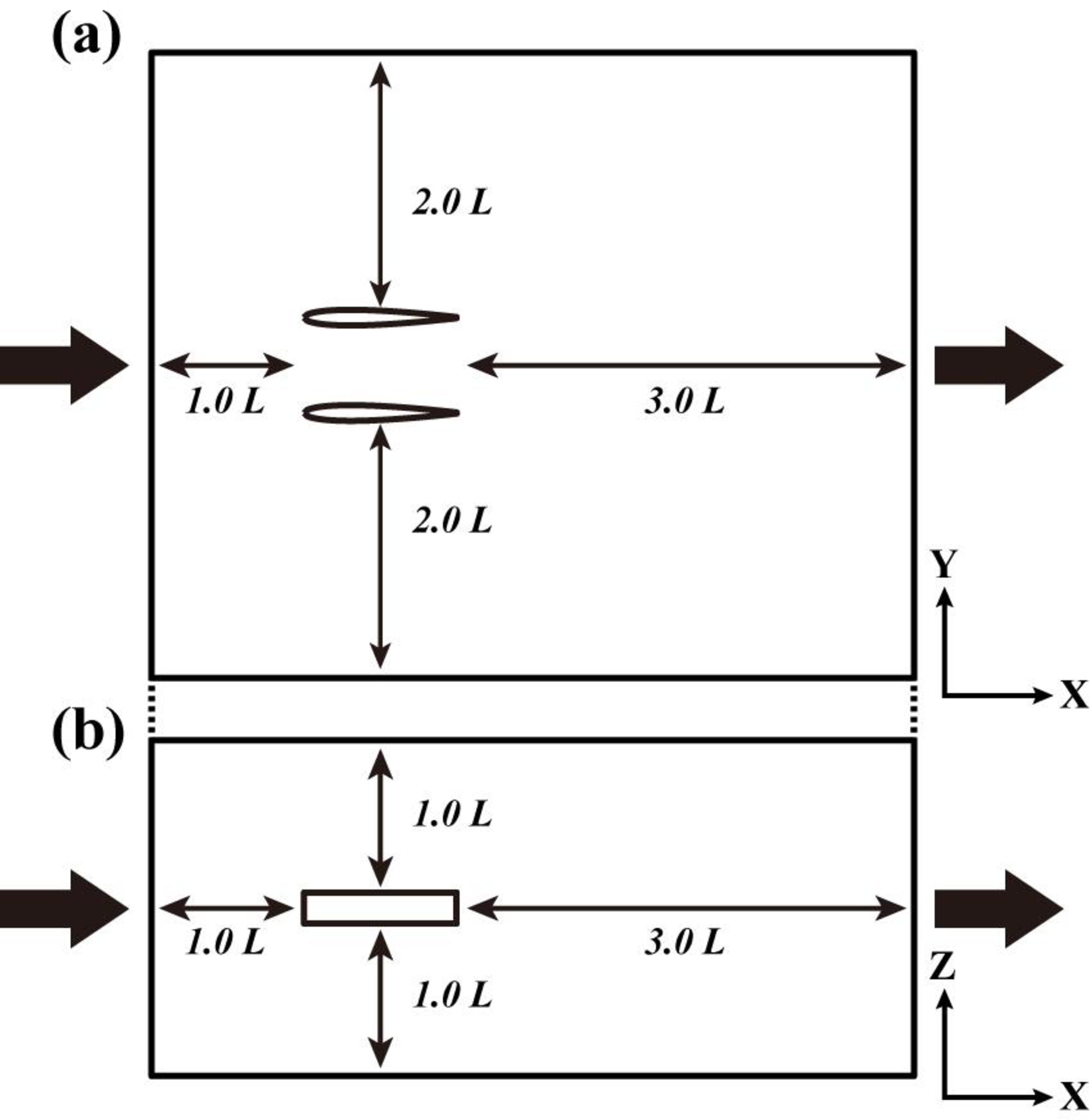
図5.パラレルスイミングのCFD解析のための解析領域。(a) 平面図,(b) 側面図。
-

図6. パラレルスイミングのCFD解析におけるメッシュ分割
-
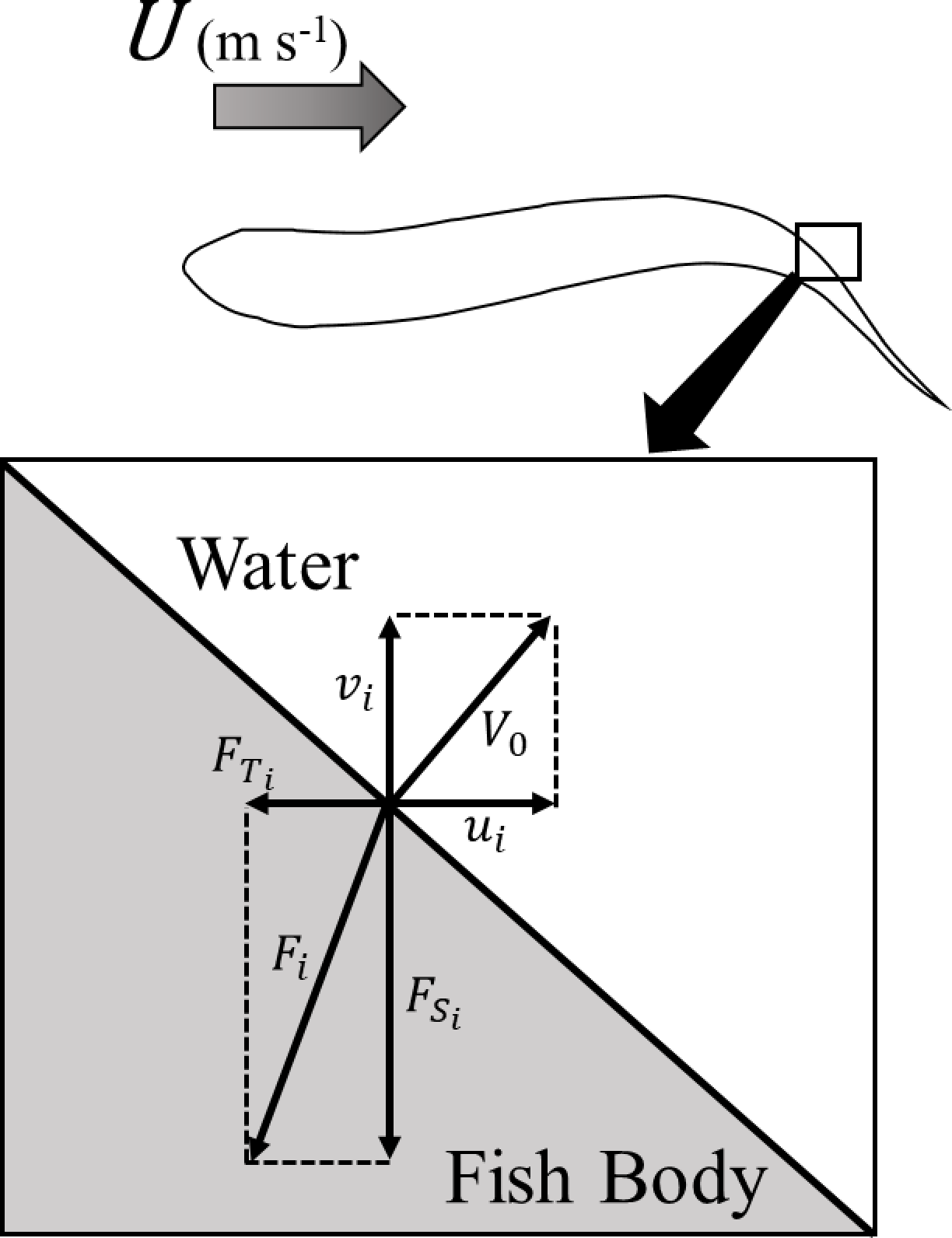
図7. 魚体表面の要素にかかる力Fiの分解
Uは流入流速、V0は要素の速度、uiは要素の進行方向速度、viは要素の横方向速度、FTiは要素に作用する推力、FSiは要素に作用する横力である。
-
-
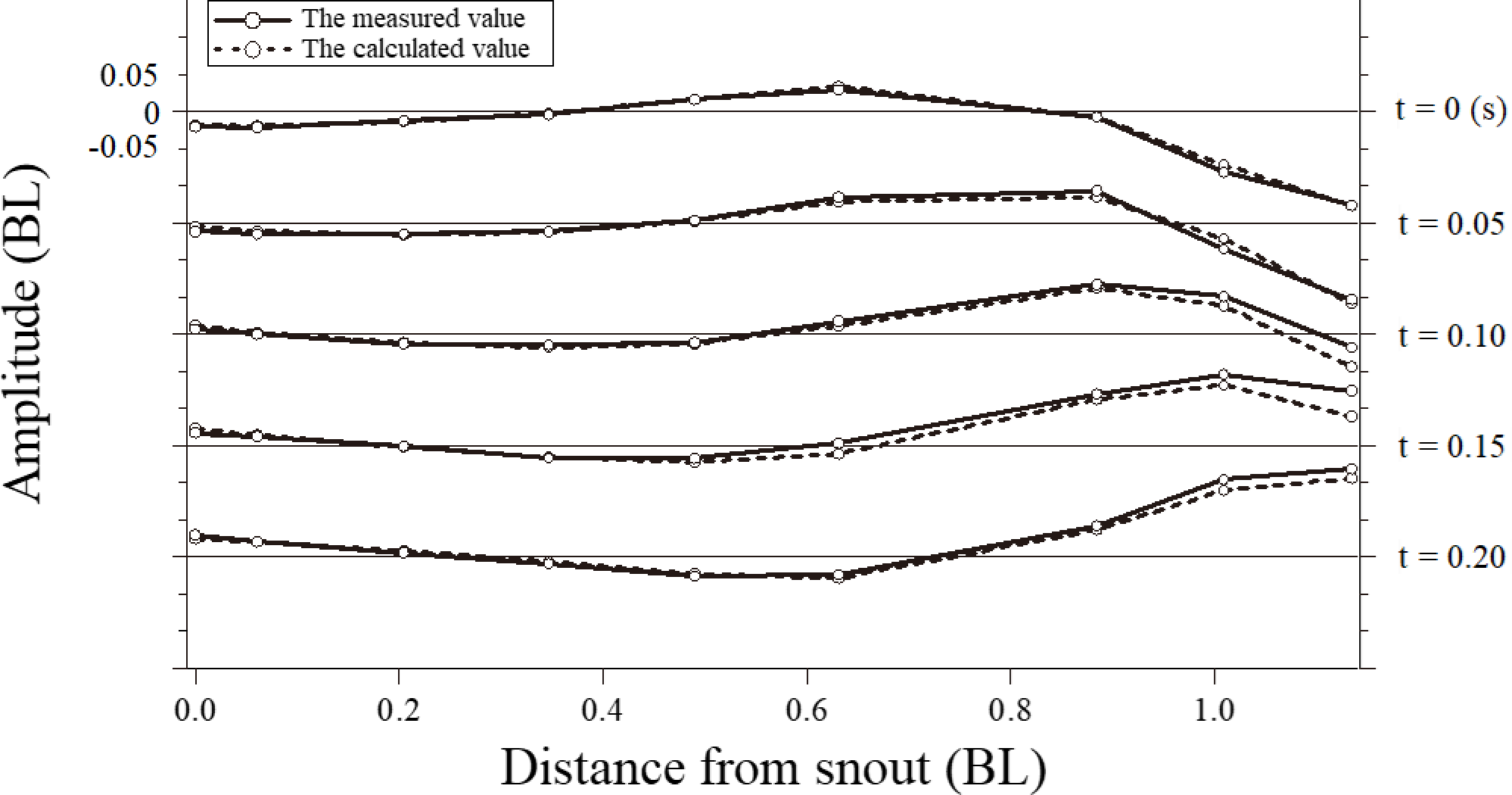
図8. 魚の体軸上の各点におけるデータの時系列
遊泳速度1.52BL s-1における実測データ(実線)と遊泳運動関数から求めた結果(破線)を比較したものである。
-
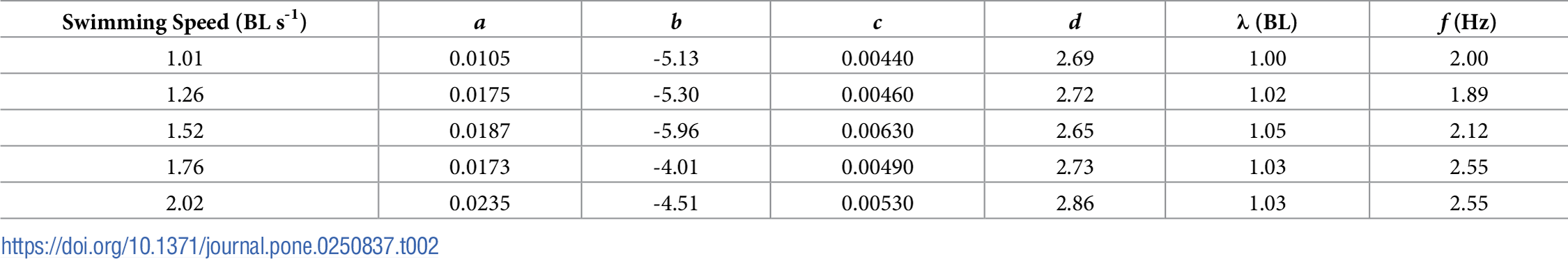
表2. 式2の定数a, b, c, dと式3の尾びれの波長(λ)と周波数(f)
-
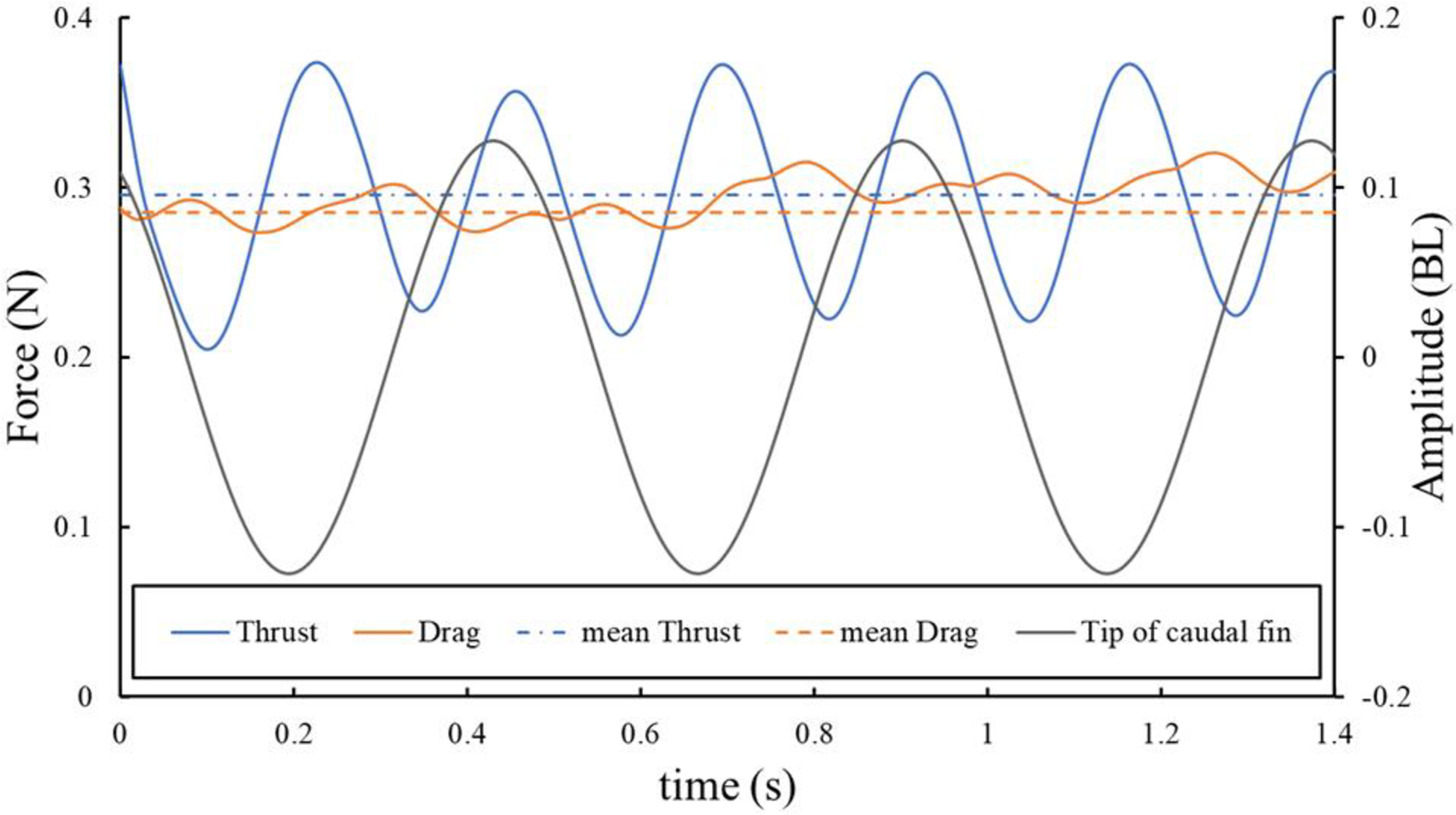
図9. 魚のモデル表面に作用する推力と抗力
これらの力は、遊泳速度1.52BL s-1で尾びれを3ストローク分行った際に作用したものである。
-
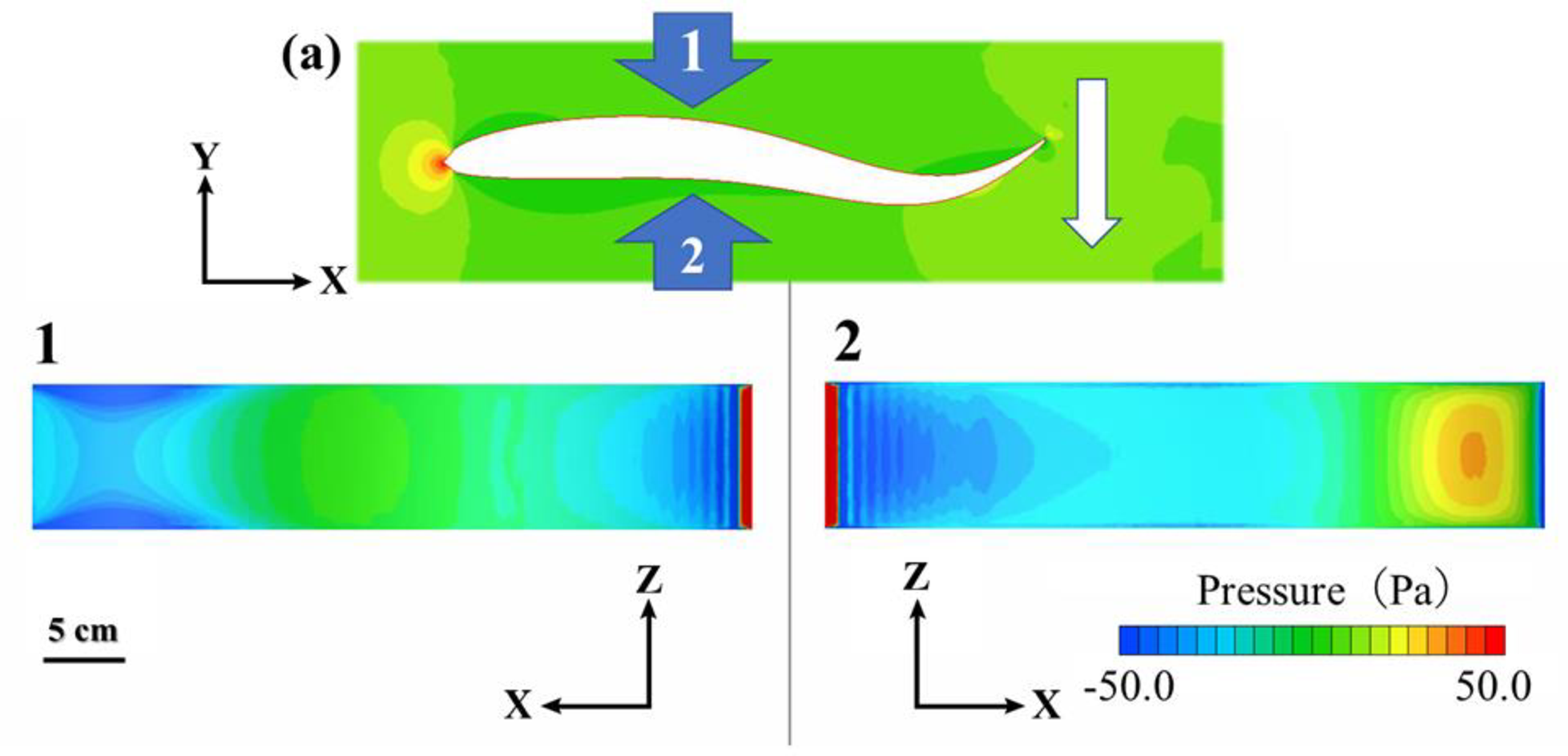
図10. モデルの面圧分布
遊泳速度1.52BL s-1における最大速度(a)時の尾びれ尾端部の右側(1)と左側(2)を示している
-
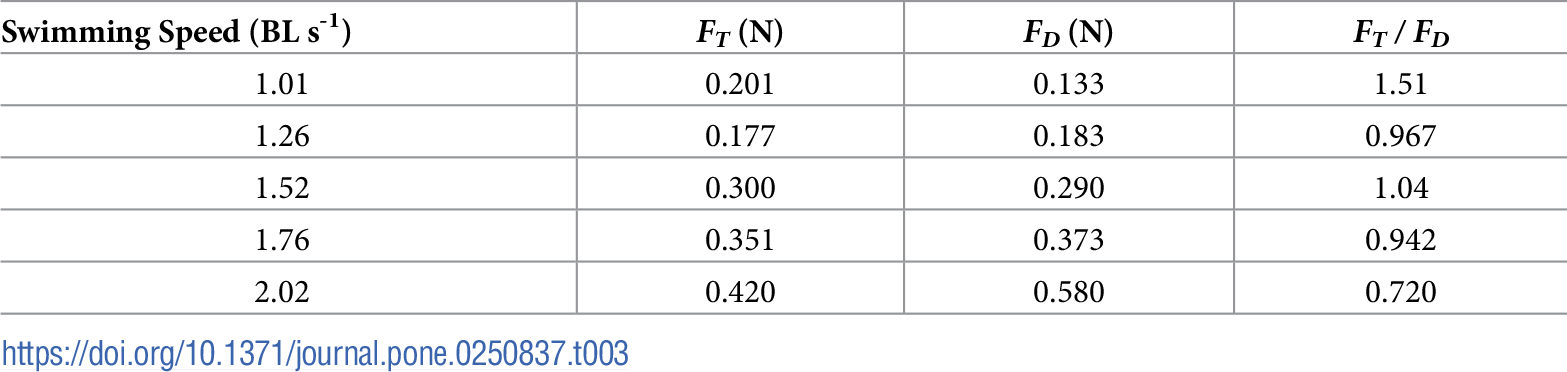
表3. 推力と抗力(それぞれFTとFDとする)の時間平均値を各水泳速度で比較した
-
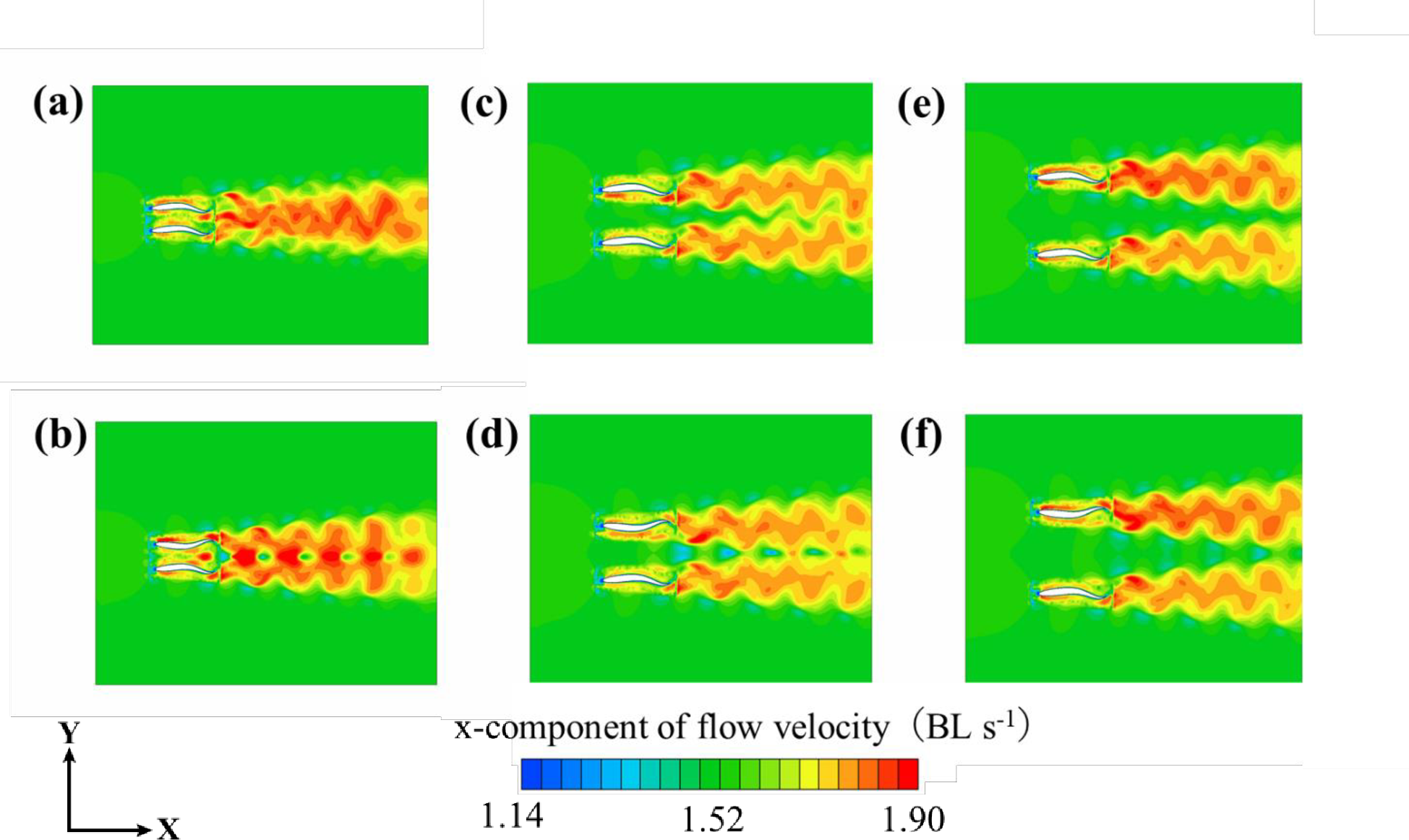
図11.モデル周辺の速度分布:(a) 0.4Lと同位相の個別距離、(b) 0.4Lと逆位相の個別距離、(c) 0.8Lと同位相の個別距離、 (d) 0.8Lと逆位相の個別距離、 (e) 1.2Lと同位相の個別距離、(f)1.2Lと逆位相の個別距離。
-
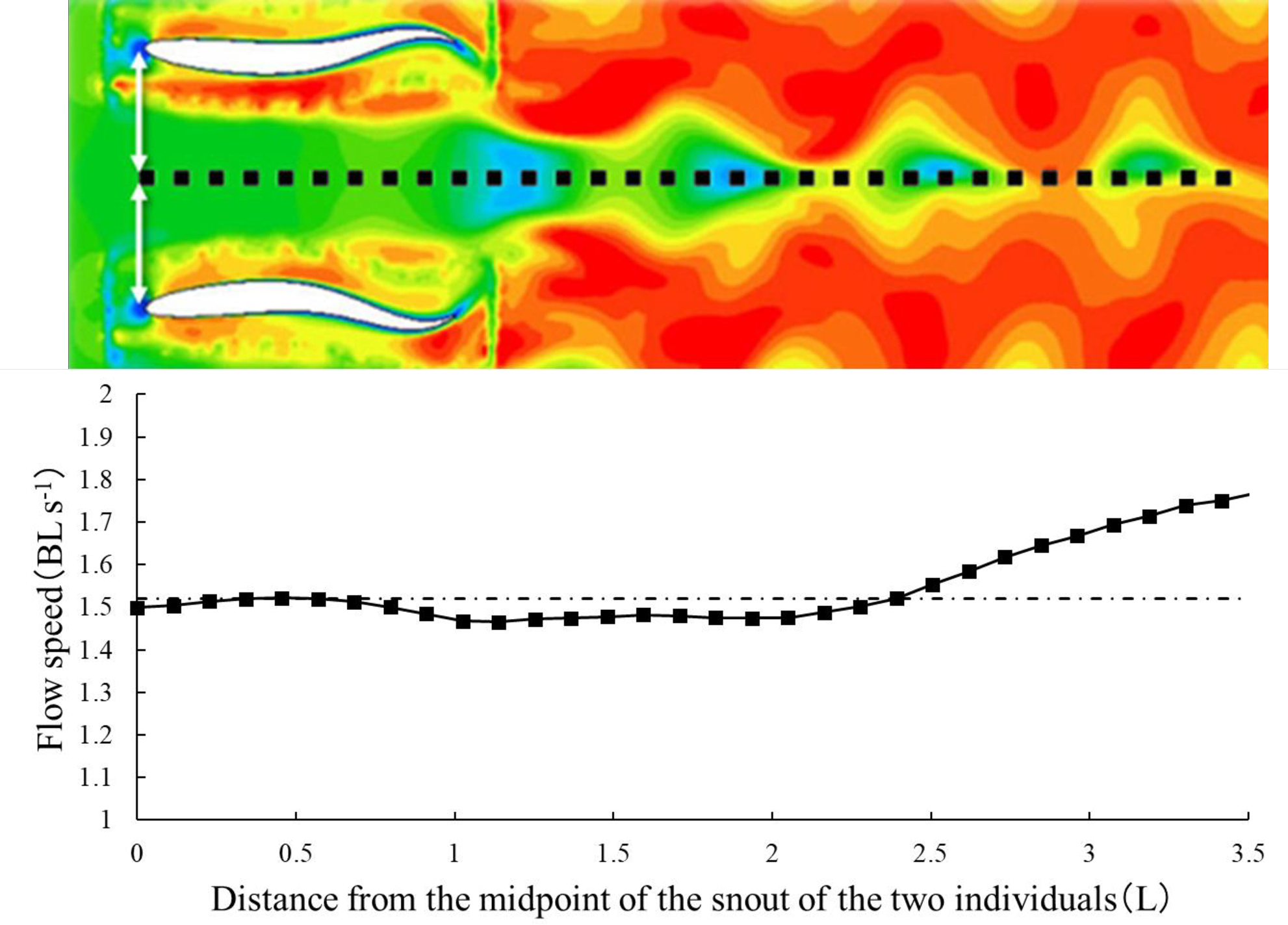
図12. 速度分布の各点における速度値
横軸は個体間距離0.8L、逆位相の条件で2個体の鼻の中点を原点としてx軸に平行な点線上の距離を表している。縦軸は点線上の速度値を表す。長破線は入口流速(1.52 BL s-1)を示す。
-
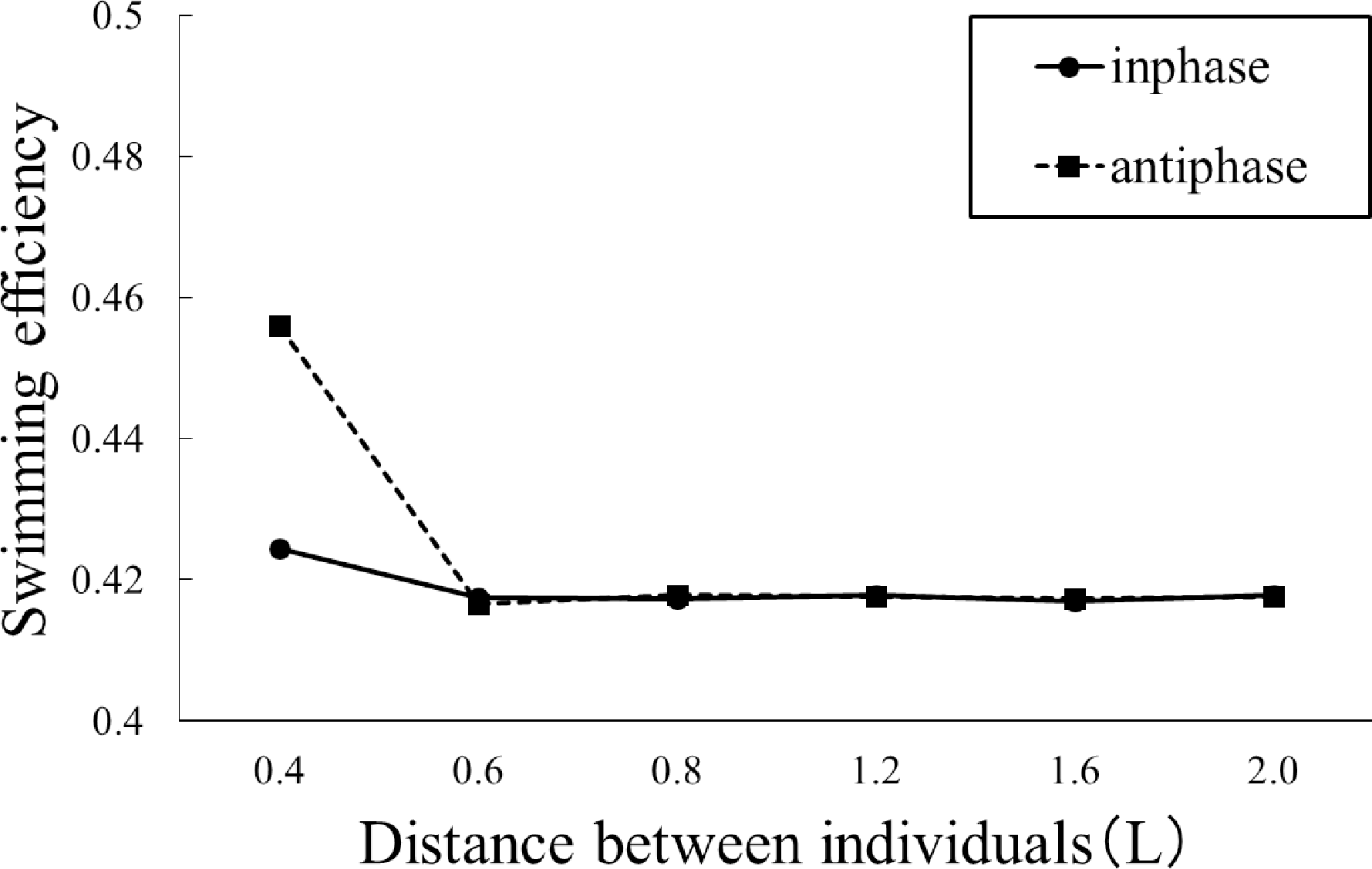
図13.同位相(実線)と逆位相(破線)の条件下で2個体が並行して泳ぐ場合の遊泳効率
-
-
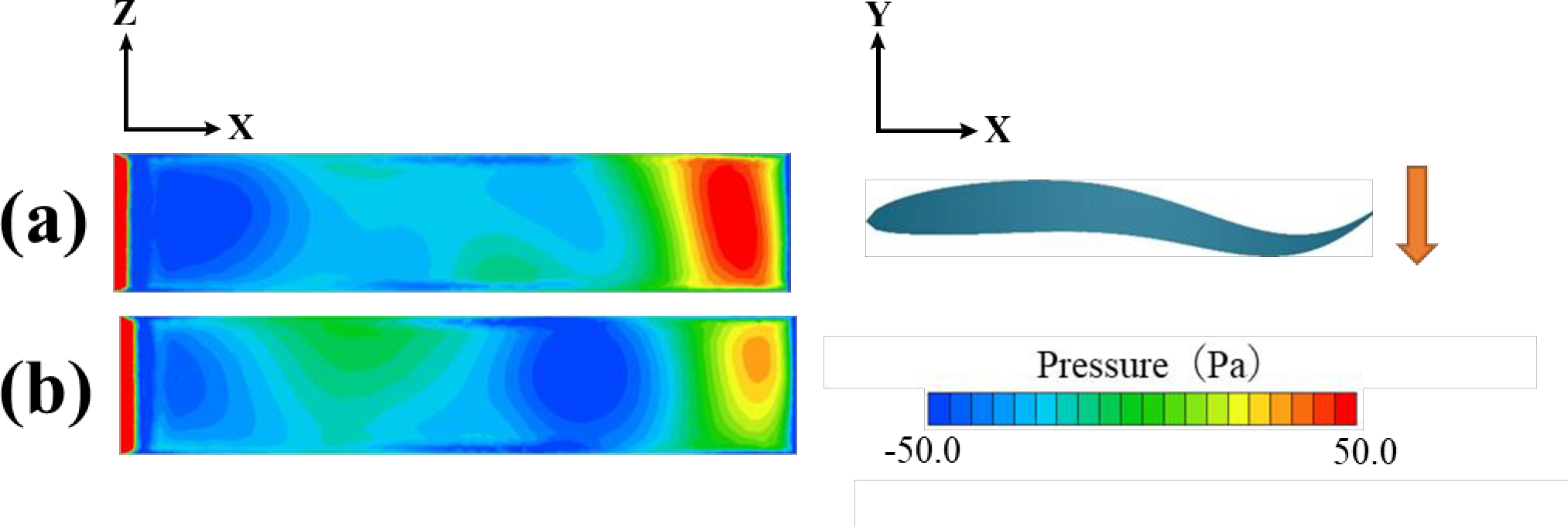
図14.尾びれの振幅運動の最大速度における表面圧力の等高線プロット:(a) 個体間距離0.4L、(b) 個体間距離2.0Lの場合。




