
クラゲ
Section outline
-
-
無脊椎動物の筋肉タンパク質(トロポニン-トロポミオシン系など)を研究してきましたが、
6年前、クラゲ好きの学生が進学してきたことを契機に、クラゲのタンパク質についての生化学的研究を始めました。
クラゲはとても魅力的な研究対象です。
●単純な動物に見えるが、未解明なことばかり
●種にもよるが、どんどんクローン増殖し、飼育や維持が容易
タンパク質の研究に関心のある人、クラゲ好きの人、当研究室で研究してみませんか?
コースの記述はやや内容が偏っていますが、ご意見や指摘事項、質問等がございましたら、tanaka-h@fish.hokudai.ac.jpまでご連絡ください。
-
-


クラゲは、原始的な無脊椎動物の刺胞動物に分類されます。なぜ原始的といえるのでしょうか。原始的とは言っても、その体には様々な機能があるのです。クラゲの体の仕組みを知ることで、生物進化の面白さに触れることができます。生物の進化、そして多様性の面白さを多くの人に知ってもらうことで、海の豊かさを守ること(SDGs14)につながればよいと考えています。
国連では2021年からの10年間を、「海洋科学の10年」に定めて、SDGsに貢献することを目指しています。
-
-
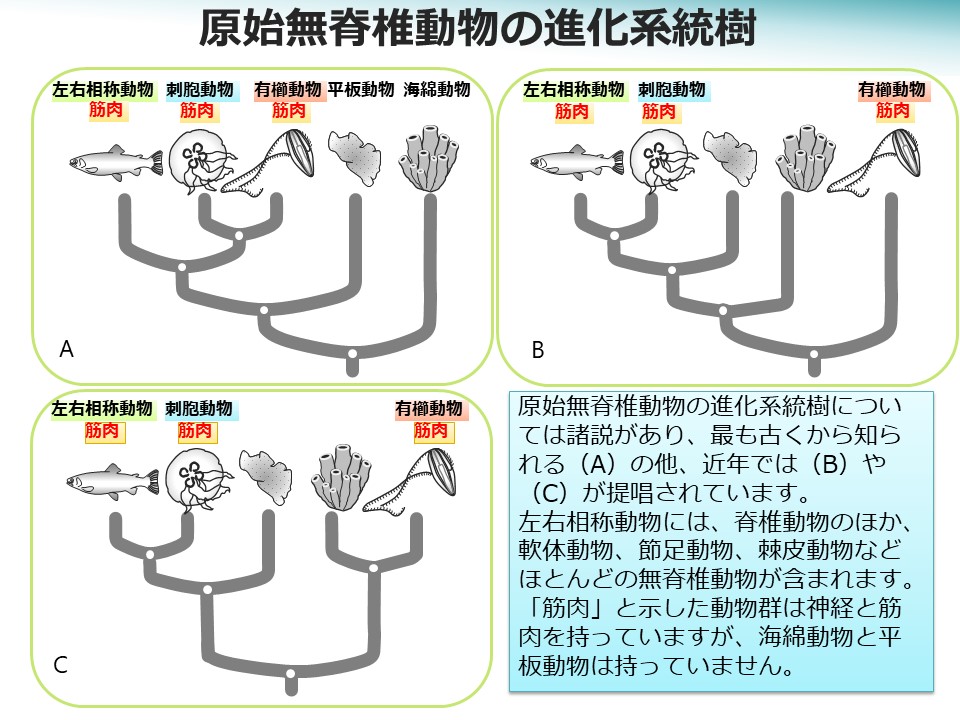
クラゲには、ミズクラゲ、タコクラゲ、オワンクラゲなど刺胞動物門(サンゴ、イソギンチャク、ヒドラなども含まれる)に属するものと、ウリクラゲなど有櫛動物門に属するものがあります。これらは古くは腔腸動物門としてひとまとめにされていましたが、昨今の遺伝子解析の成果もあって、かなり異なる動物群であることがわかってきました(下図のBやC)。
また、クラゲは約6億年前、神経と筋肉を最初に獲得した動物であると考えられています。神経と筋肉を獲得した動物は、大型化しても「行動」することが可能になり、様々な環境に適応して劇的に進化し、知性を発達させました。クラゲの筋肉は、原始的とはいっても、細胞内のエネルギー(ATP)を効率よく使って大きな力を発生させるアクチン-ミオシン系によって構成されます。また、神経には、光や接触などの刺激を受容して伝達する感覚神経、筋収縮を制御する運動神経、各種器官の働きを調節する自律神経に相当する役割分担が既に見られます。
クラゲの神経は散在神経系であり、脳はありません。しかし、観察していると、あたかも考えて行動しているような動作も見られます(動画:シロクラゲ・ポリプの捕食行動)。刺胞動物のクラゲにおいて、拍動のリズムを作り出すのは傘の縁にある神経細胞です。また、傘の縁には光や重力を感じる感覚器があり、種によってはレンズや網膜を備えた眼を持っています。そのため、傘の縁には、リング状に神経細胞が集中しており、中枢神経のように働いていると考えられています。
刺胞動物と左右相称動物の間では、筋肉や神経が動作する分子レベルの仕組みに共通点が多いのですが、有櫛動物はかなり異なる様で、神経伝達物質も他の動物とは異なっています。
-
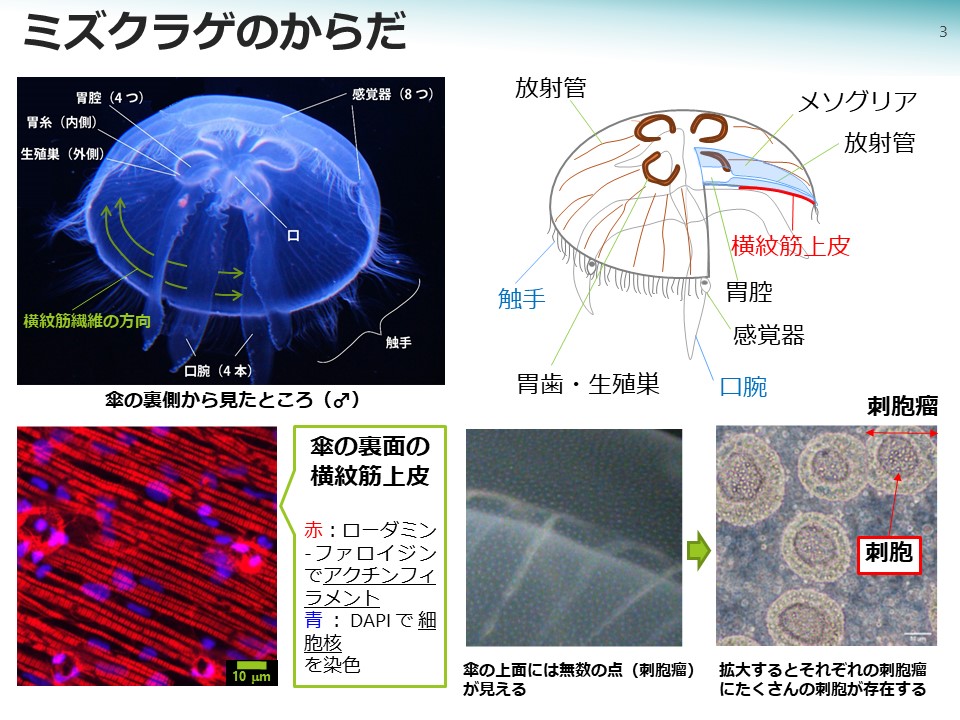
ミズクラゲの体の大部分を占めるのは水分(95%)に富んだコラーゲンのゲル(メソグリア)です。メソグリアにはアメーバ細胞がまばらに存在し、これは生体防御やゲルの修復、栄養の輸送などを担っているようです。メソグリアの表面は一層か二層の薄い細胞層が覆っていて、その中に神経と筋肉、消化と吸収、生殖細胞の形成など主要な生命機能が集約されています。
傘の縁には餌を捕らえる触手があり、触手の表面には毒針の収められたカプセルである「刺胞」がびっしりと並んでいます。餌となる動物プランクトンが刺胞に接触すると、毒針が勢いよく射出され、獲物の体に突き刺さって毒液が注入されます。仕留めた獲物は体表から分泌される粘液に絡め取られて、体表の繊毛の運動によって傘の縁に集められます。集められた餌は、さらに口腕によってなめとられ、口腕の中を通って傘の下面の中央にある口へ運ばれます。口のすぐ上は、多くの場合4つある胃腔になっていて、胃腔の外縁をかたどるように胃糸と生殖巣があるため、透明な体の中央に四つ葉のクローバーのような形が見えます。胃糸は密生する短い毛束の様に見え、引っかかった餌は分泌される消化酵素によって消化されます。胃腔が6つある個体もよく見られ、3、5、または7つの場合もあります。それらの場合は口腕もそれぞれ6、3、5、7本となります。
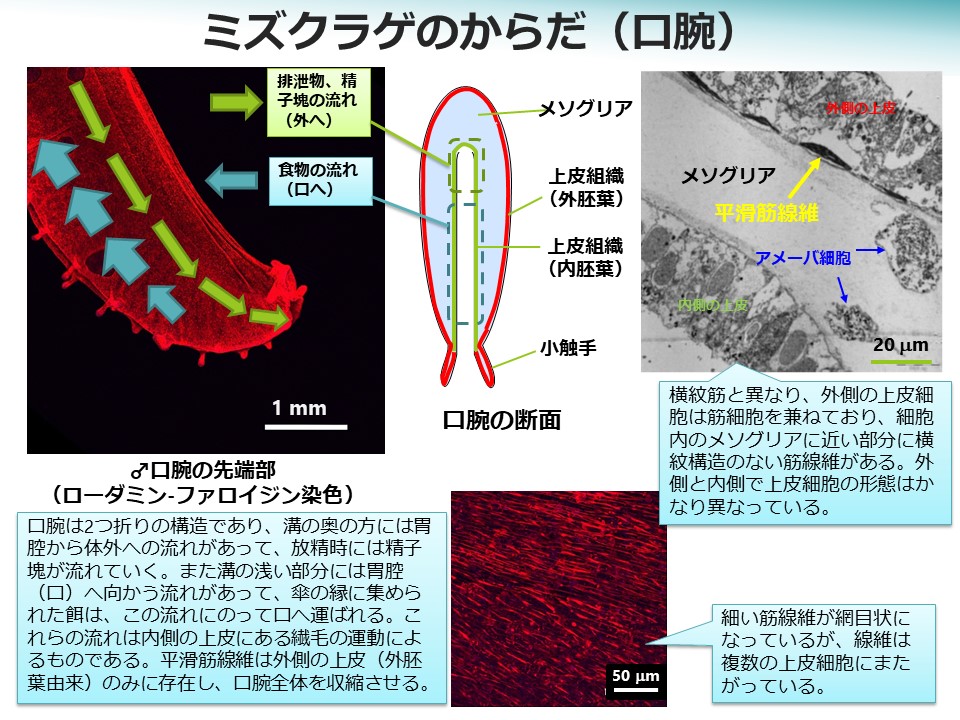
消化されて液状になった餌は胃腔から放射状に拡がる放射管を通って全身に運ばれます。放射管は腸管のように栄養吸収器官として働き、また、血管のように全身に栄養を巡らせる役割があります。ただ、心臓に相当するものはなく、放射管中の液の流れは内側に生えた繊毛の運動によるものです。胃腔から傘の縁に向かう流れの放射管と、傘の縁から胃腔に戻る流れの放射管があります。胃腔に戻ってくる液体には老廃物や消化吸収しきれなかったものが含まれていると考えられますが、それら(排泄物)は再び口腕を通って海に放出されます。口腕の中で食物が通るルートと排泄物が通るルートは異なっています。
-
クラゲの傘の拍動は、傘の裏側の上皮に存在する筋肉の収縮と弛緩によるものです。ミズクラゲの傘の裏側には同心円状に走る横紋筋の線維があって、それが収縮すると傘はすぼまった形になり、弛緩すると、傘の上方の分厚いメソグリアの弾力によって元の開いた形へ復元します。これを繰り返すことで水中を泳ぐことができます。ミズクラゲの場合、エフィラ幼生の時期には、同心円状の環状筋に加え放射状に走る放射筋を持っていますが、放射筋は成長と共に退化します。一方、キタユウレイクラゲなどでは成熟しても放射筋が残存します。拍動の周期は、成長につれて長くなり(ゆっくりになる)、水温によっても変化します。個体差もあって、傘を盛んに拍動させる個体は、拍動のたびに傘の縁の触手が良くたなびいて、たくさんの餌を集めて食べるので良く成長します。
筋繊維は1本ずつ筋細胞に収められています。筋繊維はアクチンフィラメントとミオシンフィラメントを含んでいて、ミオシンフィラメントの主成分であるタンパク質のミオシンは、ATPを分解しながらアクチンフィラメントに結合してたぐり寄せ、アクチンフィラメントとミオシンフィラメントが互いに滑り合うことで筋繊維は収縮します。ミズクラゲの横紋筋には、左右相称動物の横紋筋に似た横紋構造が観察されますが、幾つか異なる特徴もあります。一つは横紋の繰り返し単位であるサルコメアの長さが多くの左右相称動物(2.5マイクロメートル)に比べて短い(1.5マイクロメートル)という点です。同じ長さの筋繊維を短いサルコメアがたくさん連なって構成する場合と、長いサルコメアが小数連なって構成する場合を仮定すると、アクチンフィラメントとミオシンフィラメントが同じ速度で滑り合っても、短いサルコメアの筋繊維の方が全体の短縮速度は大きくなります。すなわち、ミズクラゲの横紋筋はアクチンフィラメントとミオシンフィラメントが滑り合う速度が小さくても、筋肉全体は素早く収縮できる仕組みになっていると考えられます。もう一つの特徴は、Z線の形態で、左右相称動物では電子顕微鏡観察した場合、とても濃く見えますが、クラゲの場合、不連続で脆弱な印象を与えます。

一方、触手や口腕には平滑筋があります。口腕は二つ折りになっていて、内側を食物、排泄物、卵、精子、間違えて捕らえた食べられないもの等が移動します。それぞれが通るルートは異なっていて、精子は二つ折りの溝の一番奥を通って精巣から口腕の先端まで送られ海に放出されますが、卵は口に近い部分で溝から出て行きます。これらの移動は内側の上皮にある繊毛の運動によるものです。内側の上皮と外側の上皮は細胞の形態がかなり異なっていて、平滑筋の線維は外側の上皮細胞の中にあります。これは上皮細胞と筋細胞が別々になっている横紋筋とは大きく異なる点です。平滑筋の線維は口腕全体の中で網目状につながって見えますが、実際には個々の上皮細胞に分割されて存在しています。
-
刺胞動物を特徴付ける「刺胞」は毒針の収められたカプセルであり、非常に精緻な機械的構造を有しています。刺胞は獲物を捕らえたり、外敵から身を守ったりするためだけでなく、基盤への付着などにも用いられます。有櫛動物のクラゲは刺胞を持ちませんが、代わりに獲物を粘着させて捕獲する「膠胞」を持つ場合があります。また、一部の有櫛動物や軟体動物は捕食した刺胞動物の刺胞を体表に配置して防御に役立てることが知られています。刺胞の存在はクラゲに「刺される」原因であり、多くの人々の関心を集めてきました。しかし、刺胞から毒針が射出される分子レベルの機構は未だによくわかっていません。
刺胞そのものは生きた細胞ではなく、刺細胞内に作られてエキソサイトーシスによって細胞外に押し出される構造物です。ミズクラゲの場合、体表のいたるところに刺胞が存在しますが、胃腔の内側などにも見られます。また、触手に最も多く存在しますが、口腕や傘の上面にもたくさん存在しています。意外にも傘の裏面には比較的少ないです。 -
刺胞動物は有性生殖と無性生殖の両方を行う特異な生活環を持っています。ミズクラゲの場合、成体(メデューサ)に精子と卵ができ有性生殖を行います。受精卵から発生したプラヌラ幼生はやがてポリプとなりますが、このポリプが分裂や出芽、ストロビレーションなど様々な様式で無性生殖(クローン増殖)を行い、個体数を爆発的に増やすことができます。ポリプはまた、飢餓に耐え、芽胞のようなものをつくって休眠したり、ムカゴのような細胞塊を放出して増殖することも知られています(1)。
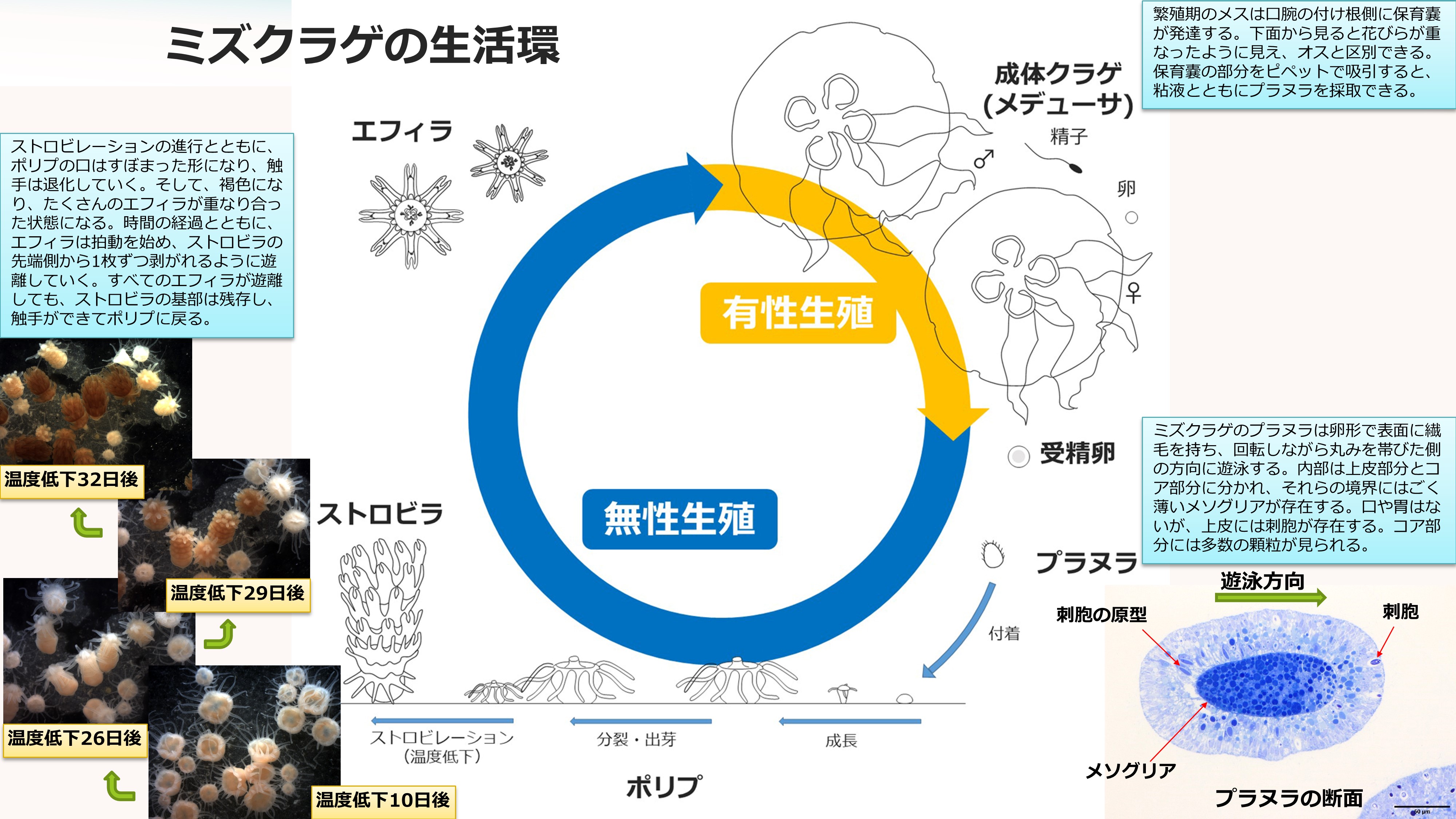
ミズクラゲは雌雄異体であり、成体(メデューサ)が成熟するとオスの生殖巣(精巣)には精子、メスの生殖巣(卵巣)には卵がつくられます。精子は精巣から口腕を通って、口腕の先端付近から放出(放精)されますが、糸状または粥状の塊(精子塊)になっています。この精子塊を顕微鏡で観察すると、たくさんの精子が尾部で束ねられていたり、絡み合ったりして、ゆらゆらと揺れており、単独で元気に泳ぎ回る精子はなぜか観察されません(画像:ミズクラゲの精子)。受精は体内受精と考えられていて、メスはこの精子塊をおそらく口腕を介して胃腔に取り入れ、卵巣で受精が行われます。このような繁殖を効率的に行うためなのか、ミズクラゲは夏期に密集することがあります(画像:ミズクラゲの密集)。受精卵は発生途上でメスの体外に排出されますが、プラヌラ幼生となるまで口腕の付け根側にある保育嚢に保持されます。保育嚢の発達したメスは口腕の形態がオスと大きく異なるので、繁殖期の雌雄鑑別は容易です。
ミズクラゲのプラヌラは、長径約0.2mmの卵形で、全身に生えた繊毛を動かして回転しながら遊泳します(動画:キタミズクラゲのプラヌラ)。プラヌラには口がなく何も食べませんが、刺胞を持っています。メスから遊離して数日後には基盤に付着し小さなポリプに変態するのですが、刺胞は付着に必要であったり、変態後に備えて体内に生産しているのでしょう。初期のポリプは触手が4本ですが、餌を捕って成長するにつれ、8本、16本と増えていきます。
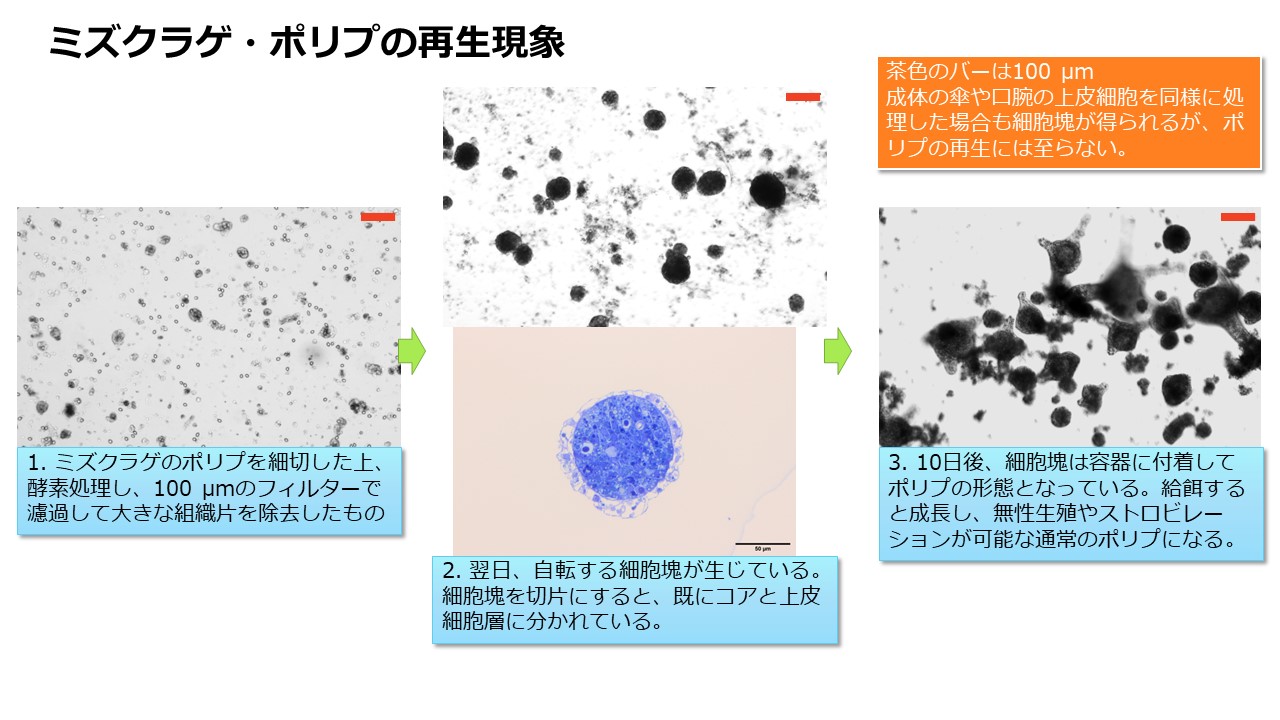
ポリプは口の周囲に16本の触手を持った小さな(<2 mm)イソギンチャクのような形をしています。付着生活をしますが、分裂や出芽によってクローンをつくり無限に増殖します。出芽は触手に似た走根(ストロン)の先端部に小さなポリプが形成される現象です。走根が基盤に付着し、そこに黄土色をしたポドシストを形成することもあります。ポドシストからは条件が整うと小さなポリプが発生します。ポリプは高い再生能力を持ち、触手の破片から個体全体が再生することが報告されています(2)。また、ポリプを細かく刻み、コラゲナーゼ等で処理して細胞レベルにまでバラバラにした懸濁液も、放置しておくと、翌日には細胞同士が集合して自転する球形の細胞塊が形成され(動画:自転する細胞塊)、1週間後には多数の微小なポリプが発生します。さらに、ポリプは数ヶ月の絶食にも耐えられます。まるで不死身ですが、環境の変化には弱く、淡水にさらせば短時間で死滅してしまいます。
季節がかわり、水温が8~10℃低下すると、ミズクラゲのポリプはストロビレーションと呼ばれる第二の変態を始めます。はじめに触手のすぐ下にくびれが1本でき、ついで、その下にさらにくびれができて、くびれの数が増えていきます。ポリプの体は次第に長く伸び、くびれは5~17本くらいまで増加します。次第にくびれは波打った形になり、色が濃くなってエフィラ幼生の縁弁ができあがります。温度低下から5週間程度たつと、先端側からエフィラが拍動を始め、1匹ずつ遊離します(動画:ミズクラゲのストロビレーション)。
(1) New observations on the asexual reproduction of Aurelia aurita (Cnidaria, Scyphozoa) with comments on its life cycle and adaptive significance. Alejandro A Vagelli, Invertebrate Zoology, 4, 111-127 (2007)
(2) Scyphistoma Regeneration From Isolated Tentacles in Aurelia Aurita. Georgia E. Laurie-Lesh and R. Corriel, J. Mar. Biol. Ass. U. K. 53, 885-894, (1973)



