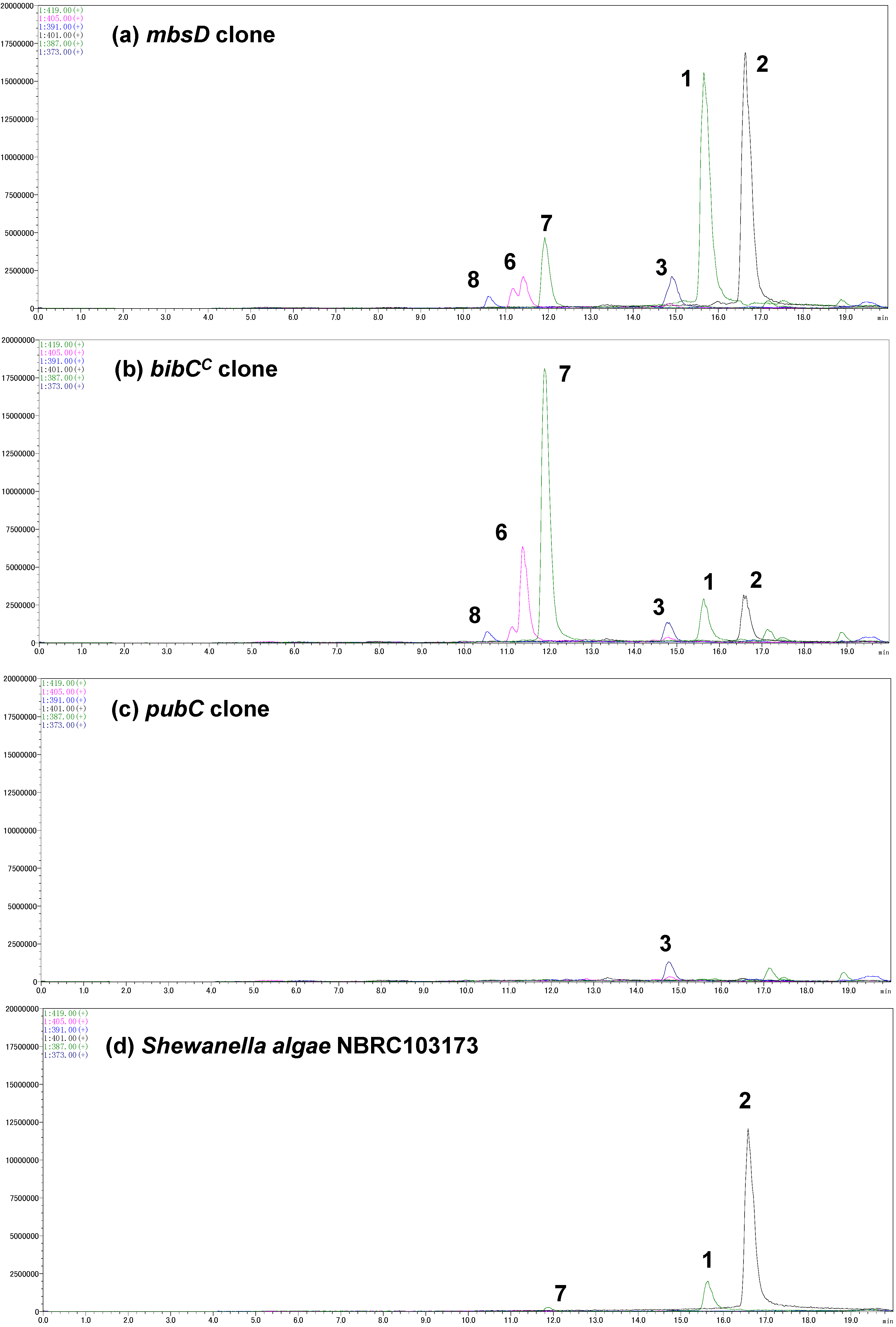
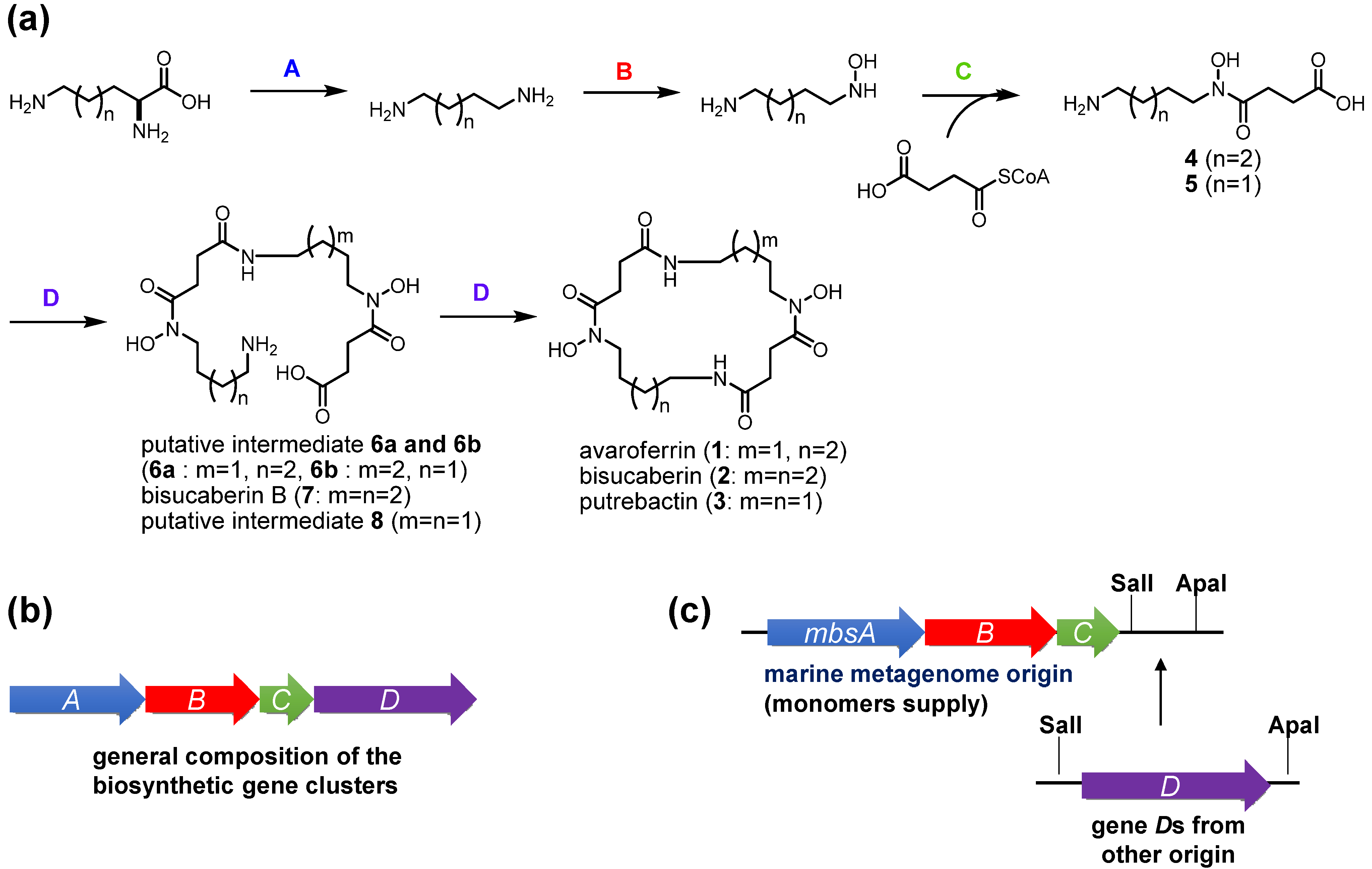
コドンを最適化したbibCCとpubC遺伝子を化学合成し、カセット法を用いてmbsCの下流に挿入しました(図4c)。これらの融合遺伝子クラスターの形質転換体を培養し、その上清をLC-MS分析に供しました(図3b,c)。bibCCを含むクローンは、化合物1と2、および少量の3を2:2:1の割合で生産しました。しかし、このクローンの主要な代謝産物は、非環状中間体(化合物6〜8)でした。特に、bisucaberin B(7)の蓄積量は、大環状化合物である2の6倍でした。これらの結果から、bibCCはアバロフェリン(1)やプトレバクチン(3)も生産できるが、その大環状化活性は、少なくとも異種宿主大腸菌ではMbsDほど強力でないことが示唆されました。pubCクローンの場合、最も多く生産されるsiderophoreはプトレバクチン(3)であり、未知のバックグラウンドピークを除けば、他の代謝物はLC-MSクロマトグラムで検出可能な量ではありませんでした。この場合の全体のsiderophore生産量はmbsDやbibCCクローンのそれと比べて3%程度であり、PubCが本来持つ機能が異種宿主で発現していないことが示唆されました。
また、同じくアバロフェリン(1)生産菌であるShewanella algae NBRC103173株のsiderophoreも解析しました(図3d)。興味深いことに、この S. algae 株は主にbisucaberin(2)と少量のアバロフェリン(1)を生産し、検出可能な量の 3 を生産しませんでした。この生産パターンは、化合物 1-3 を 1:2:1 の割合で生産する S. algae 株 B516 の報告とはかなり異なっていた。培養条件は完全に同じではないが、生産プロファイルの大きな違いから判断して、これらの酵素セットは同一種内でも代謝の多様性を持たせる鍵の一つとなり得ることが示唆されます。
検討した3つの縮合酵素、MbsD、BibCC、PubCのアミノ酸配列の同一性は約60%であるが、どのアミノ酸残基が酵素機能の違いに関与しているかは不明です。また、酵素によるオリゴマー化反応やマクロシクリル化反応の分子機構もほとんど分かっていません。これらの酵素や関連する他の生合成経路の構造生物学的な解析が進行中です。
本研究では、1つの酵素系で複数のsiderophoreを生産することを実証し、HSD系siderophore生合成酵素の驚くべき汎用性を明らかにしました。これらの酵素のユニークな特性は、競争的な微生物群における生産者にとって有利に働く可能性があります。例えば、ビブリオ属細菌の中には、siderophore特異的受容体を発現することにより、他の細菌が産生する外来siderophoreを利用するものが知られています。この現象はsiderophore piracyと呼ばれ、構造的に関連する siderophore1〜3の生態学的機能の個別の違いを説明すると思われます。V. alginolyticus B522は、特定の受容体を用いて2と3(ただし1ではない)を「盗む」ことができます(siderophore uptake system)。したがって、複数の siderophoreを生産する能力は、 siderophoreの海賊行為を回避する戦略を表している可能性があります。本研究は、 siderophore生合成遺伝子の配列をわずかに変えるだけで、酵素の性質が大きく変化し、その結果、 siderophore生産に大きな影響を与えることを実証しています。
mbsA-D metagenomic siderophore生合成遺伝子群は、S. algae B516由来の推定アバロフェリン(1)生合成遺伝子群との配列類似性から、Shewanella sp.または関連株のものと共通の祖先を持つ可能性があります。しかし、培養可能な細菌からこれまでに分離された他の関連酵素と比較して、mbsA-Dクラスターの触媒機能が明らかに異なることは、環境ゲノムの全く予期しない機能の発見をもたらすかもしれない、機能ベースのメタゲノム解析アプローチの威力を示している。