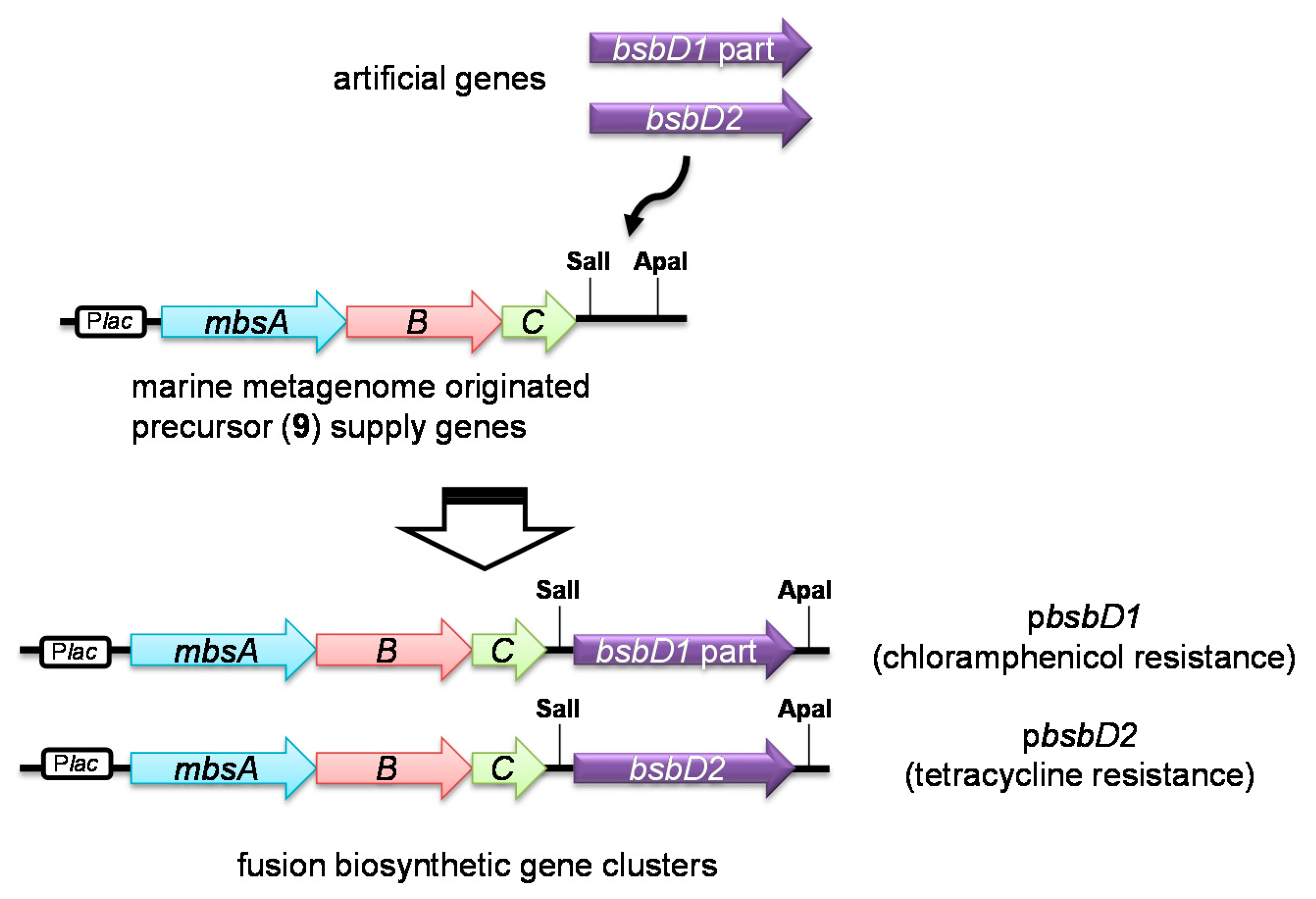
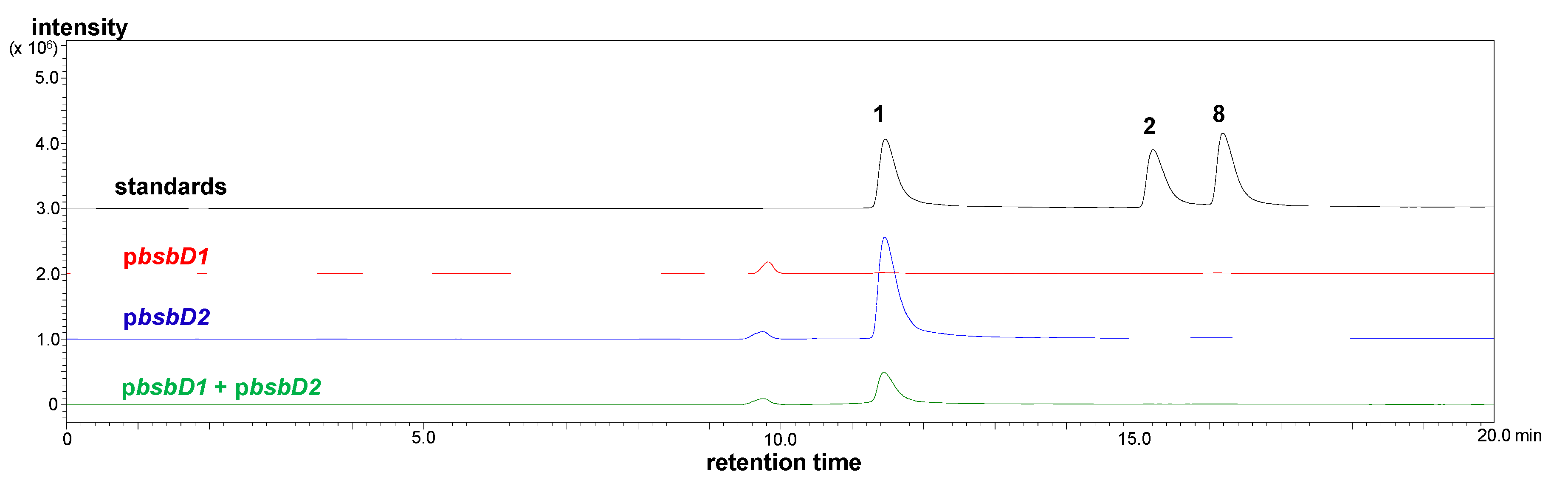
今回の実験ではBsbD1部分の機能を証明することはできなかったが、以上の結果から、本システムではbsbD1部分が効率的・機能的に発現していないか、コードされているタンパク質であるBsbD1部分が本質的に不活性であることが示唆されました。
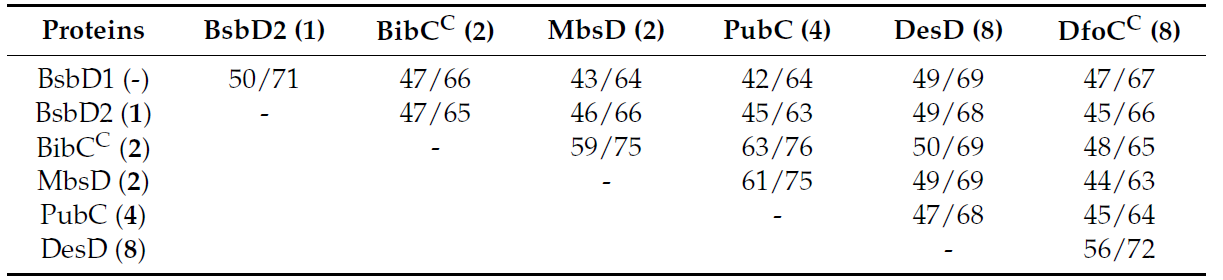
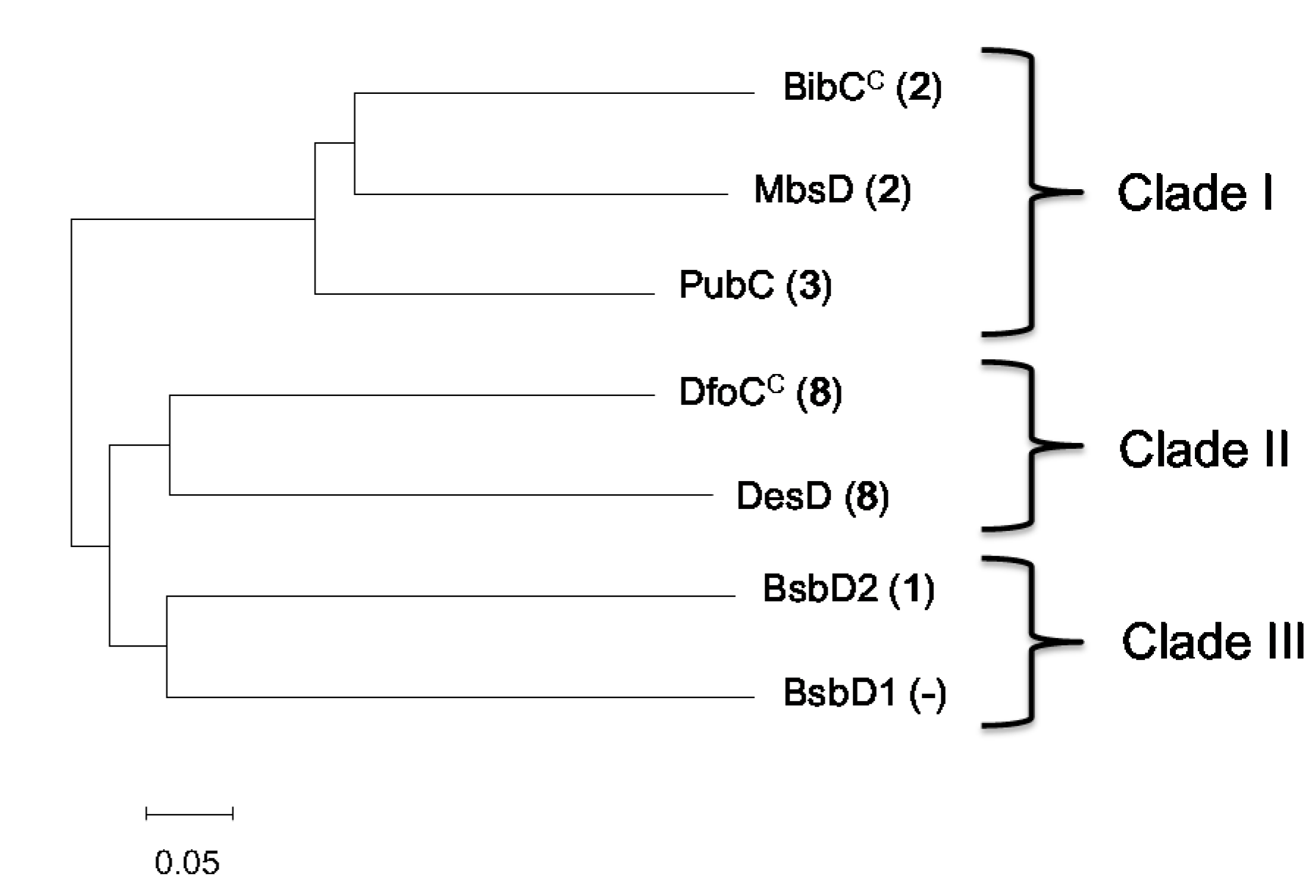
現在までに、HSD (9, 10) ベースのsiderophoreの生産を担ういくつかのアミド結合形成性大環状化酵素(酵素Ds)が、様々な細菌種から実験的に特徴付けられている(例えば、 desferrioxamine E (8) 合成酵素 DesD と DfoCC は Streptomyces coelicolor と Erwinia amylovora から、 bisucaberin (2) 合成酵素 BibCC と MbsD は Aliivibrio salmonicida と marine metagenome から、プトレバクチン (4) 合成酵素 PubC は Shewanella sp から、 アルカリギン (5) 合成酵素 AlcC はBordetella pertussisから、それぞれ同定 (表 2))。本研究では、BsbD2が大環状化能を持たない酵素Dの最初の例であることを証明しました。したがって、BsbD2の塩基配列を解析することで、このほとんど知られていない酵素の分子基盤が明らかになると期待されました。