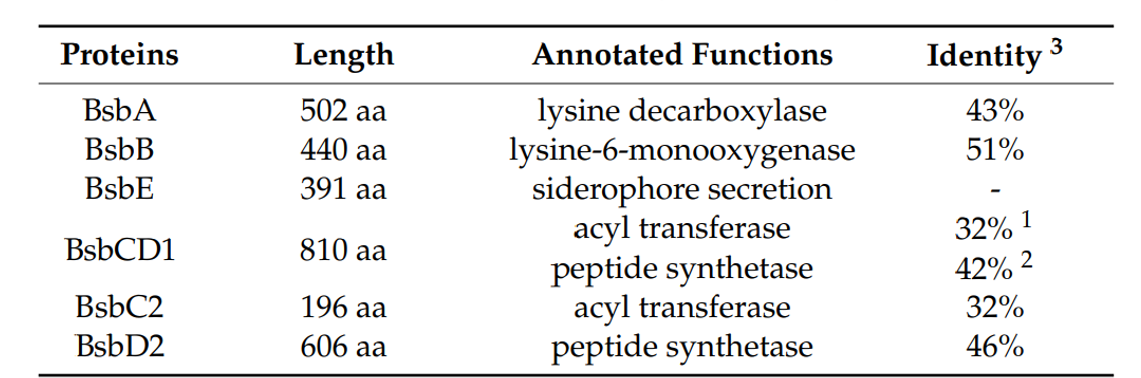
Bacteroidetes門からはHSD型siderophore生合成遺伝子は報告されていないが、多様な細菌分類群のアミド結合形成酵素(酵素Ds)の間で高度に保存されたアミノ酸配列を利用して、保存部分を縮退プライマーセットを用いたPCR増幅によりクローン作成しました。T. mesophilumのゲノムDNAから増幅したDNA断片は、既知の生合成遺伝子と高い類似性を示し、本種に類似の遺伝子クラスターが存在することが示唆されました。そこで、T. mesophilumのゲノムライブラリー(約9.6×10⁴個のフォスミドクローンから成る)をPCR増幅スクリーニング法によりスクリーニングし、全遺伝子クラスターを含むクローンを同定しました。ヒットしたクローンのショットガンシーケンスを行った結果、推定bisucaberin B(1)生合成酵素をコードする遺伝子クラスター(全長8840bp)が存在し、bsb (bisucaberin B)クラスターと名付けられました(図3、表1;受入番号LC090204)。このクラスターには、これまでに報告されている関連生合成遺伝子クラスターで一般的に見られる4つの遺伝子(A~D)ではなく、6つのオープンリーディングフレーム(ORF:bsbA, B, CD1, C2, D2, E)が含まれていました。