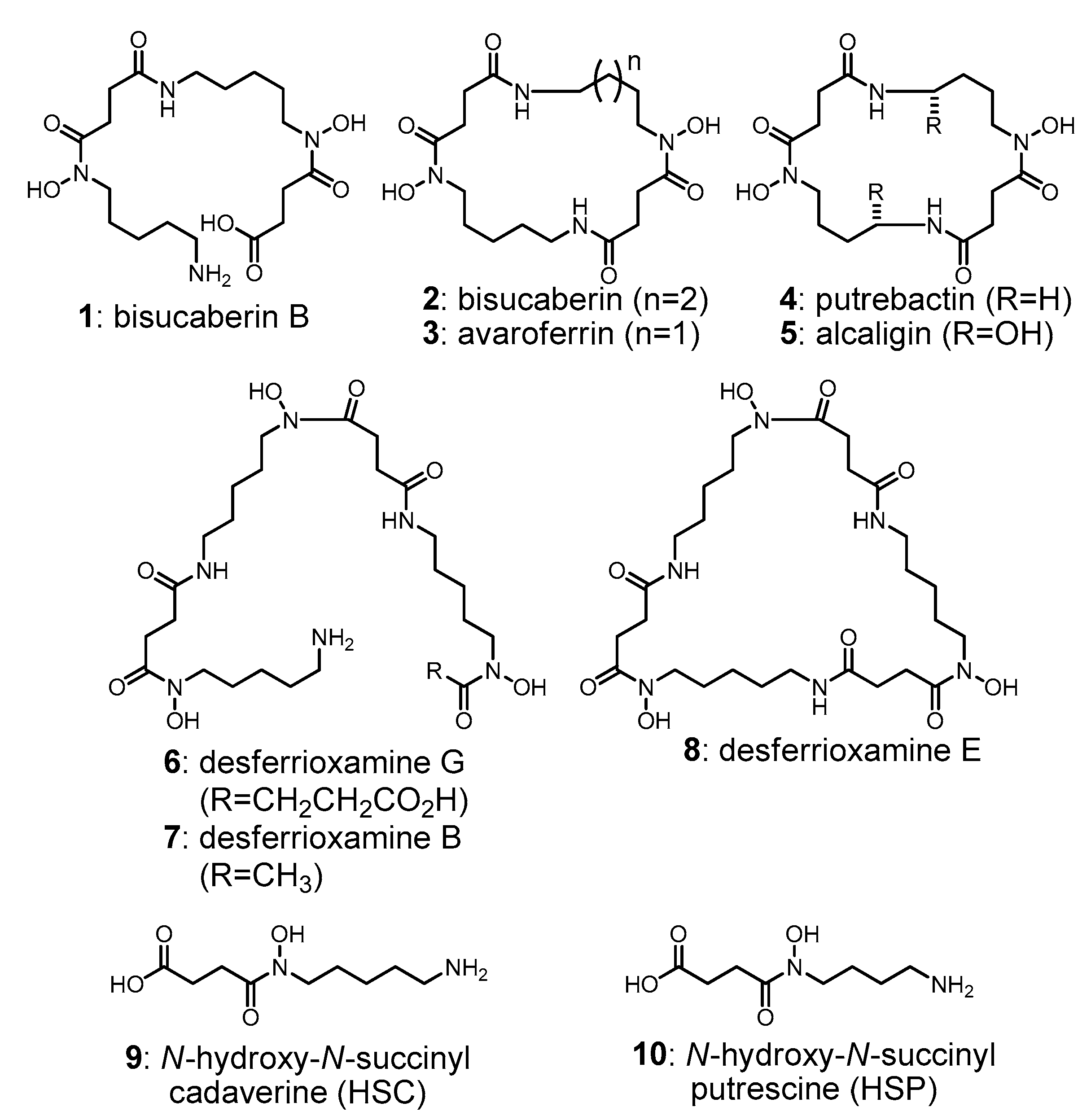
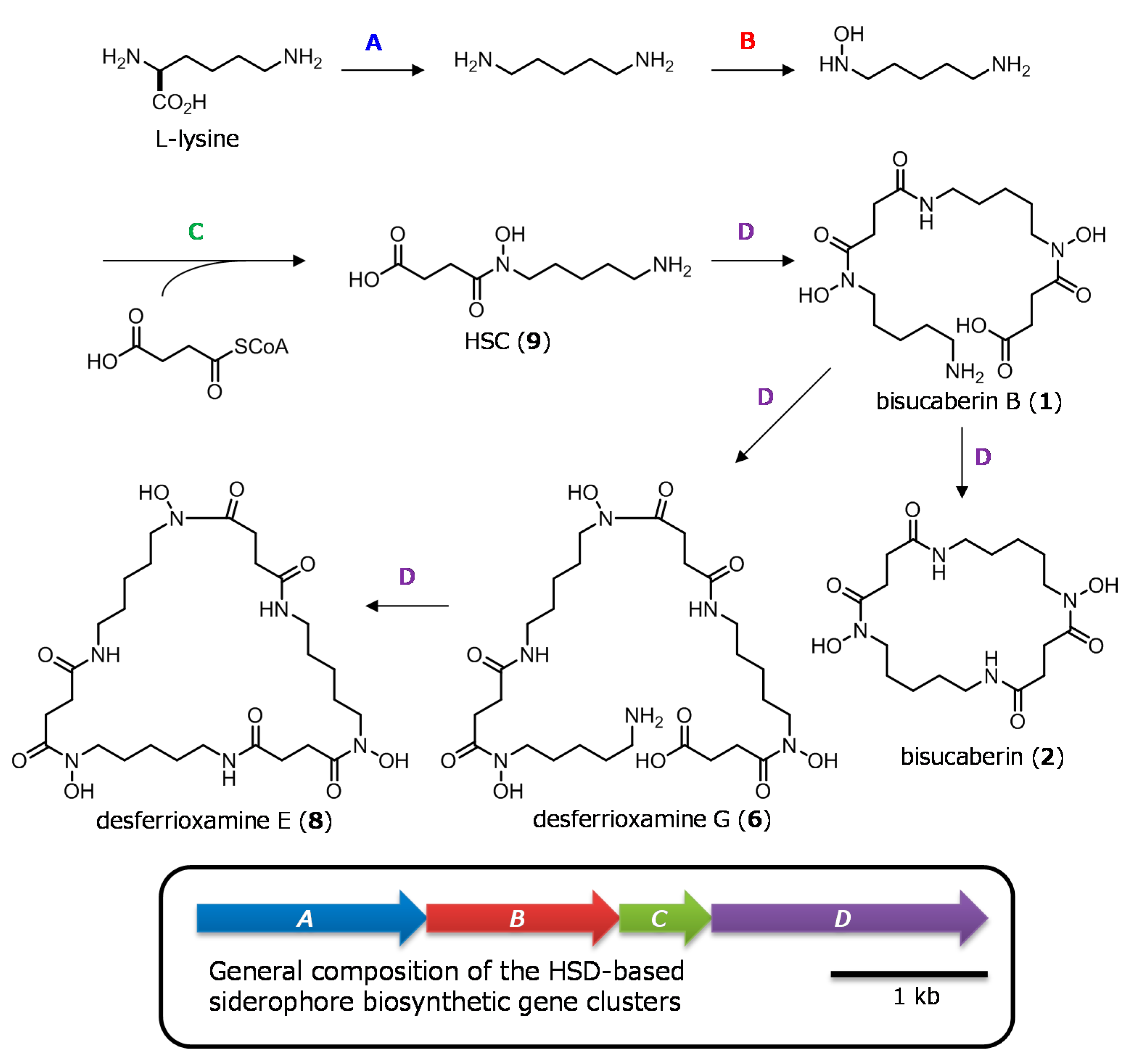
現在までに、HSD系siderophoreの生合成を担う遺伝子クラスターがいくつかクローン作成されています。これらのクラスターは一般に4つのタンパク質をコードしており、最初の3つの酵素(酵素A〜C)は、アミノ酸(リジンおよびオルニチン)から、脱炭酸、N-水酸化、 succinyl-CoAとの縮合を順次行って共通の主要中間体であるHSDs(9および10)の生成を触媒しています。第4の酵素(酵素D)は、HSDモノマー間の複数のアミド結合の形成(9, 10)を触媒し、その後のhead-to-tail環化反応によって、最終的な大環状生成物を得ます(図2)。これらのアミド結合形成酵素(酵素Ds)は、非リボソーム型ペプチド合成酵素の新しいグループを構成しています。また、酵素Dsは幅広いHSDを基質として受け入れ、多様な生理活性を有する大環状最終生成物を生成することが報告されているが、オリゴマー化反応や大環状化反応の制御の分子機構はほとんど分かっていません。