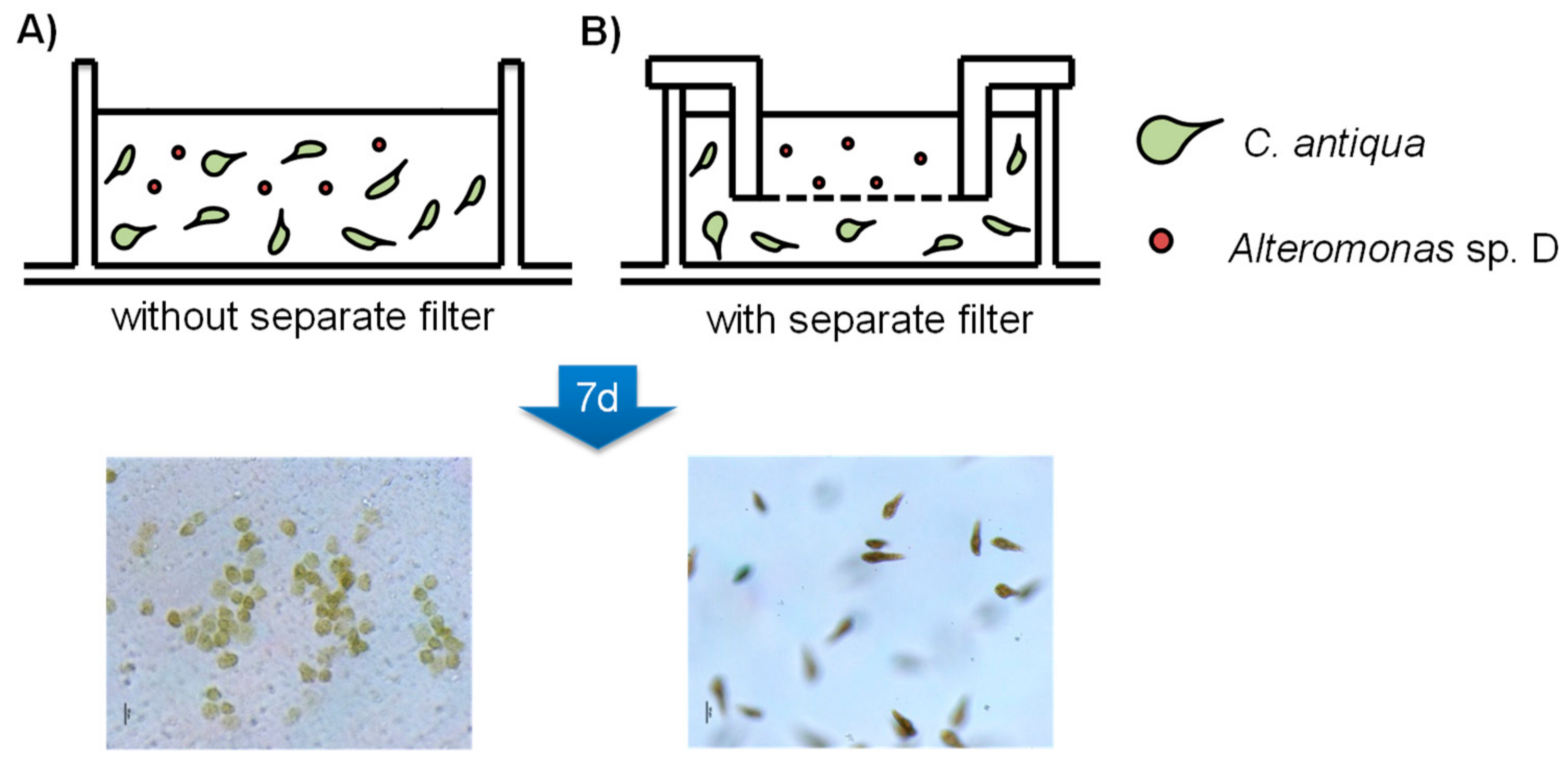
以上の実験結果から、Alteromonas sp. Dの殺藻作用の少なくとも一部は、化合物1〜4に起因することが強く示唆されたが、これらの化合物は実際の海洋環境とは全く異なる培養条件で生産されたものでした。そこで、微細藻類が繁殖する環境に近い条件下で、questiomycinの生産とその殺藻作用について検証を行いました。C. antiquaをAlteromonas sp. Dと共培養し、過剰な栄養分を含まない改良SWM-3培地で培養しました。植え付け2日後、ほとんどの藻類細胞は死滅し、フラスコの底に沈殿したが、D株を含まないC. antiquaの培養物は正常な成長を示しました。
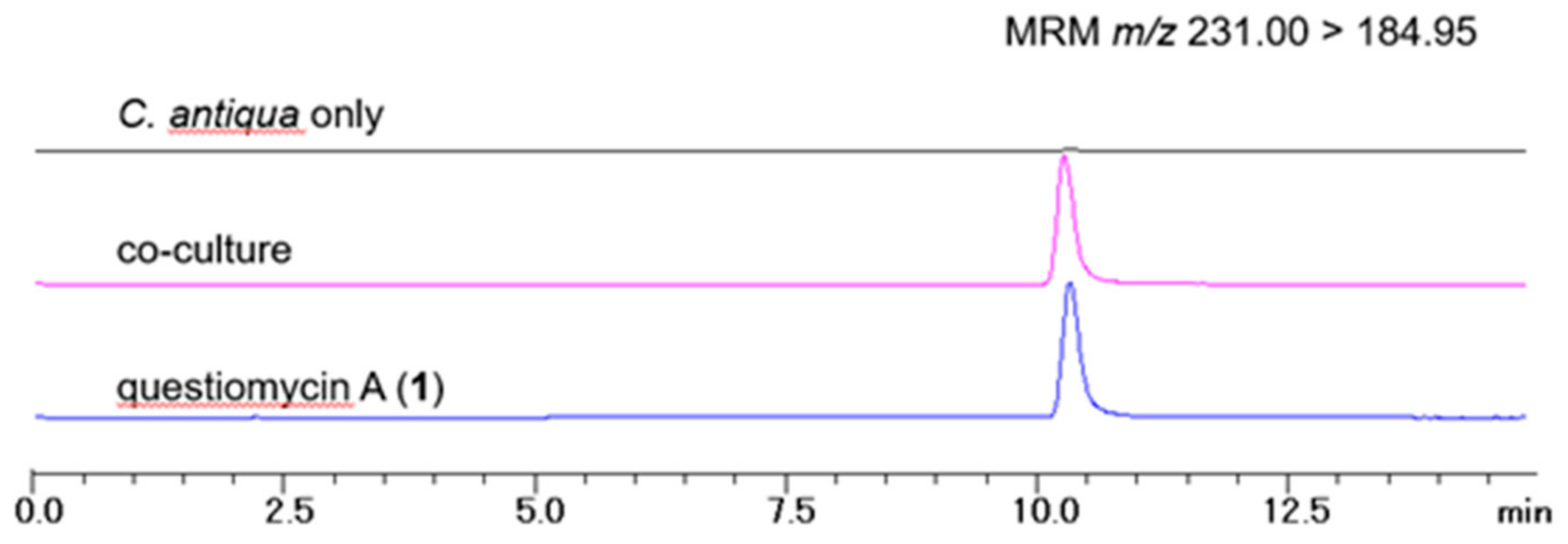
上記の共培養培地と藻類細胞の抽出物をLC-MS分析した結果、藻類が繁殖している海水と同様の条件で、確かに化合物1が生成していることが確認されました(図3)。ただし、その計算濃度は 0.31 ng/mL (1.46 nM) であり、化合物 1 の LC₅₀ 値の約 400 倍でした。なお、この実験では他のquestiomycin 2-4 は検出されていません。藻類の初期密度や培養期間が異なるため、2つの実験における1の活性を直接比較することはできないが、濃度の大きな差は、1の作用のメカニズムに何らかの洞察を与えるものであると考えられます。