
魚類発生工学
セクションアウトライン
-
-

図1.胞胚期の胚盤の移植の図。A)赤く染色した胚と未染色の透明な胚。B)染色と未染色の胚盤の上側を切断。C)切断直後。D)切断した部分を入れ換えて移植した直後。E)修復された胚盤。
-

図2.図1と同様の移植だが、一つの胚盤全体を別の胚の動物極側に移植した移植胚。下は、左から移植胚、胚盤の上部を切除した胚、無処理の胚、を示している。どれも正常に発生する。
-
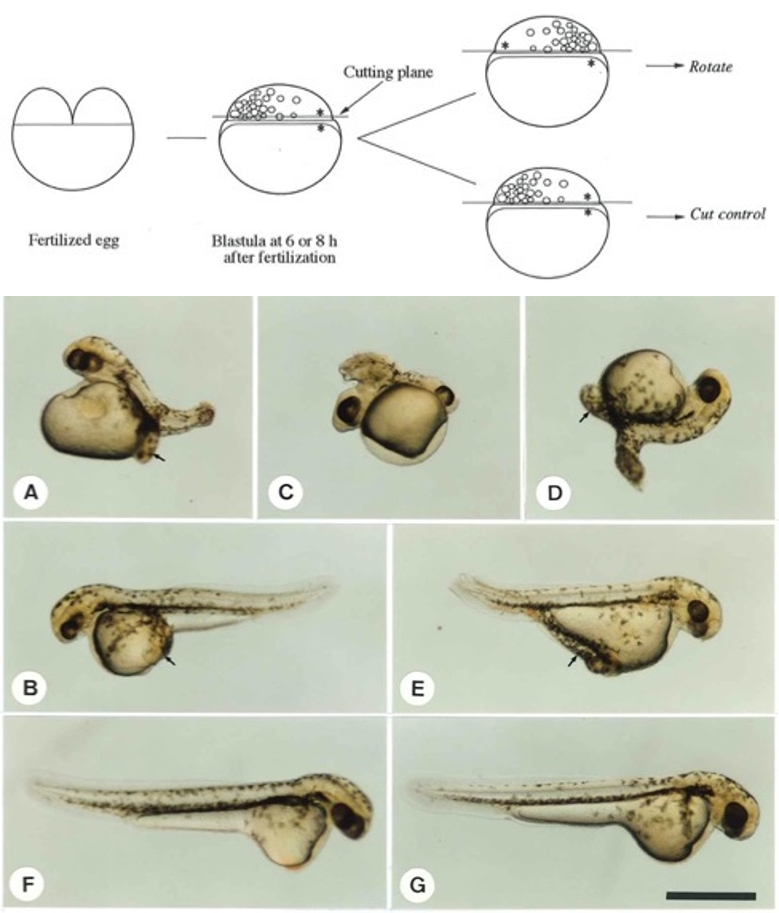
図3.A-Eは胞胚期の胚盤を卵黄細胞から切り離し、水平に180°回転させて再結合した胚。A-Dは中期胞胚期、BとEは後期胞胚期に手術を行っている。Fは360°回転させ再結合した胚、Gは無処理の胚。
-
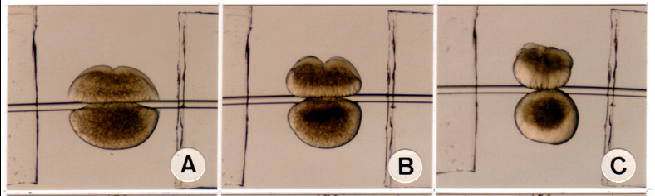
図4.2細胞期の胚を細いテグスで押し切ると、細胞質を持った動物極半球と、卵黄の多い植物極半球に切り分けられます。
-
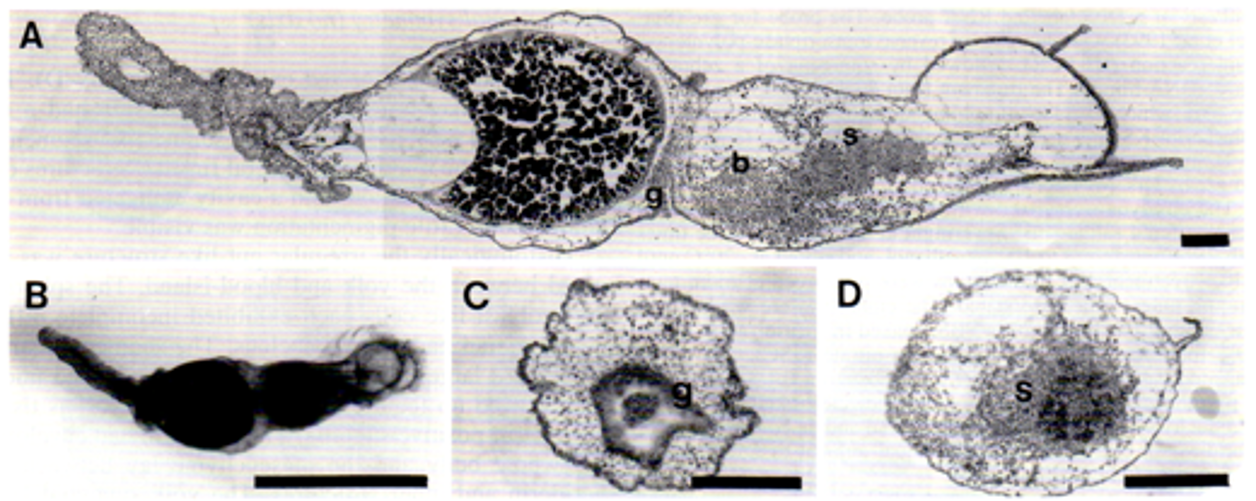
図5.初期の卵割期に動物極半球を切り出し、培養すると正常な形の胚にはならず、回転相称の胚になってしまいます。
-

図6.1細胞期に細胞質のみを切り出すと、細胞塊のみの胚(KとL)になってしまいます。
-
-

図7.初期の卵割期に細胞質内にあるvasaというmRNAを特異的に染めると、卵割溝の両側に集積しているのが分かります(aからeまで)。
-

図8. bucky ballというタンパク質に蛍光を与えると、生殖細胞質の分布を調べることができます。
-

図9.GFP蛍光で始原生殖細胞を可視化したワカサギ胚。
-
-

図10. キンギョの胚にフナの胚を移植すると、赤い色素と黒い色素を持った「キメラ個体」ができます。この個体は、キンギョの卵とフナの卵を両方産みます。ですから体細胞キメラであると同時に生殖系列キメラでもあります。
-

図11.蛍光で光らせた生殖細胞のみを移植した生殖系列キメラ個体です。蛍光を持った生殖細胞は、移植された個体の中で自律的に生殖腺へと移動します。
-
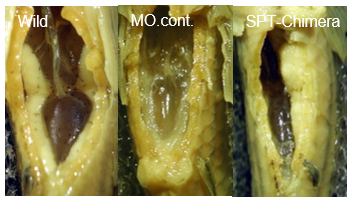
図12.左から無処理の対照個体(Wild)、PGCを作らせなくした対照個体(MO.cont)、PGCを作らせなくした個体に別の個体から一つのPGCを移植した個体(SPT-chimera)、のそれぞれの生殖腺。Wildでは一対の発達した生殖腺が認められるが、MO.contでは認められない。さらにSPT-chimeraでは一方の生殖腺のみが発達している。
-
-

図13.借り腹による種苗生産の模式図。ドナー(ここではカレイ)の受精卵からPGCを取り出し、ホスト(ここではキンギョ)の胚へ移植し、キンギョでカレイの配偶子を生産させる。
-
-

図14.染色体の数を増やすためには、細胞分裂を抑制する技術が必要です。左側のカラー写真は、サクラマスの第一卵割の図です。
-
-
-

図15.

