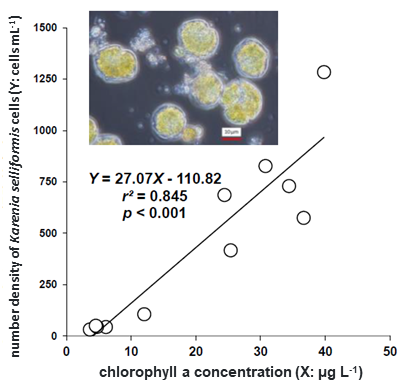
High densities of K. selliformis
were distributed at fixed sites except off Hiroo, along the Kushiro coast, and
near the shore of Akkeshi. In situ sea surface chlorophyll a concentrations (X:
µg L-1) ranged between 3.6-39.8 µg L-1 and the cell
number density of K. selliformis (Y: cells mL-1), Y = 27.07 X
-110.82, a significant positive relationship with a contribution of 85%. Using
the slope of this regression equation, the intracellular chlorophyll content of
K. selliformis was estimated to be 37 pg cell-1. A
generalized linear model analysis with cell number density of K. selliformis
as the objective variable and environmental factors as explanatory variables
revealed a positive relationship with phosphate among the various nutrients.
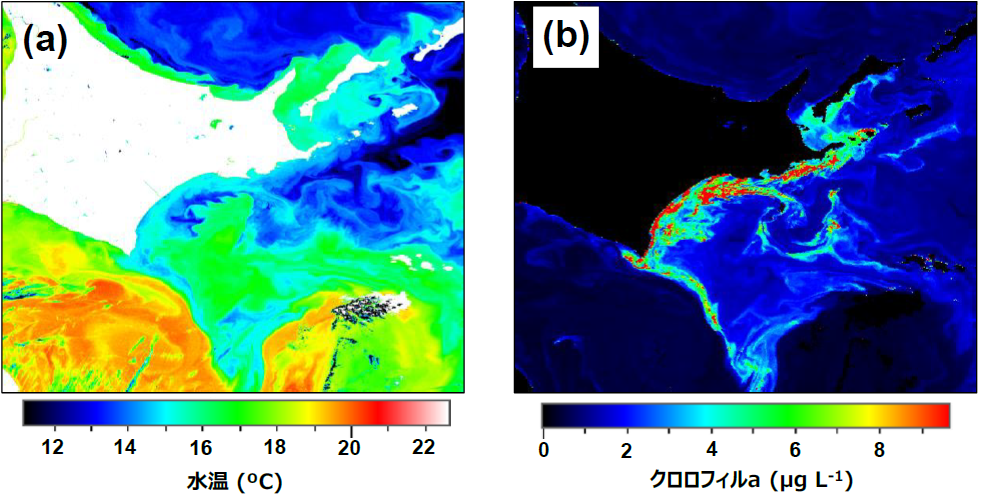
Throughout the study area, sea surface
phytoplankton densities ranged from 38-9033 cells mL-1. Cluster analysis
based on cell count densities for each species divided the phytoplankton
communities into four communities, A-D. Of the four phytoplankton communities,
18 of the 32 stationary sites were in community A, which had the highest number
of stationary sites, and community A was dominated by the dinoflagellate K.
selliformis, which accounted for 92% of cell count density, with an average
cell count density of 999 cells mL-1, outperforming the other
communities (77-152 cells mL-1). Sea surface temperatures in the
study area ranged from 13.9-18.1°C and salinities from 27.6-33.7. The sea
surface temperature and chlorophyll a concentration based on satellite data
during the survey period also indicated that high chlorophyll a concentrations
were found in the low-temperature water mass east of Cape Erimo and beyond.
Red tides have been reported to have
occurred along the east coast of Hokkaido in the fall seasons of 1972, 1983,
1985, and 1986. Although the period of the red tides varied from year to year,
they were reported to have occurred from September 3 to October 1, in the
Tokachi coast as the sea area, with dinoflagellates as the causative algae, and
with reduced catches of salmon in set nets as the damage. It is noteworthy that
whenever red tides occur in these eastern Hokkaido waters, there is always a
description that the water temperature is higher than usual.
The genus Karenia has the ability to
move by means of two flagella. Karenia mikimotoi can move vertically
around the water depth of 20 m per day at a speed of 2.2 m h-1,
while K. brevis can move at a speed of 1 m h-1. Karenia
selliformis has also been observed to have extremely high locomotion under
the microscope. The cell size of K. selliformis is about twice as large
as that of K. mikimotoi and K. brevis, suggesting that the
diurnal vertical migration capacity of K. selliformis is high.
The specific gravity of seawater becomes
lighter under high water temperature and low salinity conditions. This means
that when the sea surface is warmer than usual in the low-salinity Oyashio
region, the thermocline will develop strongly. When the water temperature
dynamic layer develops, nutrients in the shallow areas below the layer are
depleted, making it difficult for phytoplankton (diatoms, etc.), which do not
have the ability to move, to proliferate. On the other hand, dinoflagellates of
the genus Karenia, which have high mobility, so enabling diurnal
vertical migration which distributed in the surface layer during the daytime
for photosynthesis, and dive to the depths below the thermocline at night to
replenish nutrients. In 1972, 1983, 1985, and 1986, red tides are explained as
a “rainfall-type red tide” that when the water temperature was higher than
usual and the thermocline developed, only dinoflagellates with high mobility
were able to increase for a long time causing
the species composition was simple, and increased river flow after rainfall
provides nutrients to the coastal zone, which allows a single species to
proliferate and form a red tide.
Large-scale harmful red tides of K.
selliformis in 2021 were observed extensively in the open ocean, and it is
difficult to interpret them as transient “rainfall-type red tides” along the
coast. Considering these factors, the mechanism of K. selliformis red tide
in the Pacific coast of Hokkaido in the autumn of 2021 is as follows: “Seawater
temperature rises → Thermocline strengthens → Diatoms (competing organisms)
decrease → Karenia selliformis with the ability to migrate increase by
supplying nutrients through diurnal vertical migration →Surface community
dominated by K. selliformis →passage of low
pressure → weakening of stratification / vertical mixing / increase in nutrients
in the luminous layer → red tide by K. selliformis” is a possible
scenario.