
テナガの秋:負けたあいつと知らないあいつ
セクションアウトライン
-
前のコース:卒研論文化までの道のり←|目次|→次のコース: ニュージーランド滞在記
-
-
ある個体を別の個体と区別する認知能力 cognitive ability を個体識別能力と呼ぶ。脊椎・無脊椎動物を問わず,生き物が他個体と社会関係を構築する上で根幹を成すコミュニケーション能力の1 つであり,主に血縁や繁殖パートナー同士,そして敵対関係にある個体間で実証されてきた(Tibbets and Dale 2007)。個体識別が成立・継続している間は,相手を識別している個体(受信者 receiver)は「識別対象の個体(送信者 sender)が独自に持つ特徴(識別キュー recognition cue)」と「送信者に関する特異的な情報」を結び付けて学習・記憶し,将来遭遇する際にこれらの情報を利用して行動を調節する(図5.1:Tibbets and Dale 2007)。個体識別が成立すると,攻撃行動が減少することが多い。なお,キュー cue とは受信者が感知して利用する情報を指す(Maynard-Smith and Harper 2003)。
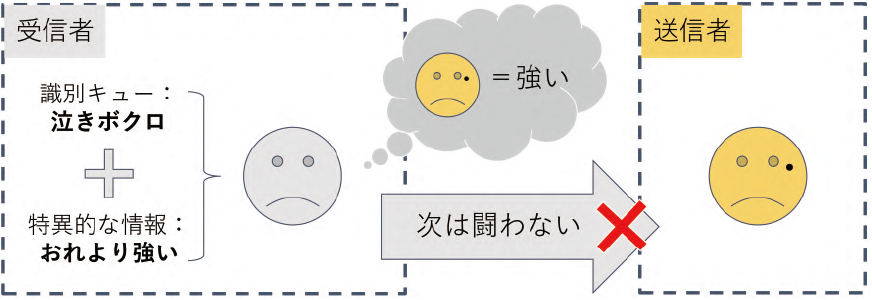
図5.1 敵対関係における個体識別のイメージ。灰色の個体は黄色の個体と過去に闘争し、敗北した。黄色の個体と再び闘争しても勝ち目が薄いのであれば、この個体との闘争は避けたほうが余計なコストを払わずに済み、有利である。このような状況では、黄色の個体独自の特徴 (識別キュー) とその個体に関する特異的な情報 (自分よりも強い) を結び付けて黄色の個体を識別し、将来の闘争を回避する個体識別能力が進化しうる。
-
識別能力は必要とされる認知能力の高さによって,「クラスレベルの識別 class-level recognition」「真の個体識別 true individual recognition」に大別されている(Gherardi et al. 2012)。「クラスレベルの識別」は,近隣の個体/見知らぬ個体,優位/劣位,繁殖パートナー/パートナーではない個体,血縁/非血縁など,主に “クラス” と呼ばれる2つの社会的なカテゴリに基づく他個体の識別である(二値的な認知 binary recognition:Wiley et al. 1991)。片や「真の個体識別」は,特定の個体を集団のあらゆる他個体から識別できる,非常に高度な識別能力である(図5.2)。
クラスレベルの識別の1 つに「既知個体の識別 familiar recognition」がある。これは,遭遇相手が “既知の個体 familiar” か,初めて遭遇した “未知の個体 unfamiliar” かを識別する個体識別能力を指す。既知個体の認知は「その個体との遭遇経験があるかないか」のみに基づく識別なので,識別した相手が属する社会カテゴリを特定する必要はない。しかし,たとえば繁殖パートナーをそのほかの個体から識別するためには,そのパートナーと以前から遭遇し,既知の関係性 familiarity を構築していることが不可欠である(つまり,相手に関する学習が必要)。そのため,既知個体の識別は,クラスレベルあるいは真の個体識別の第一段階とも考えられている(Gherardi et al. 2012)。一方で,クラスレベルの識別には,単なる雌雄の区別など,相手に対する学習が不要な識別も含まれるため,既知個体の識別を第3の独立した個体識別能力だと考える研究者もおり(Chak et al. 2015 など),学術用語としても familiar recognition は class-level recognition より一般的である。本書もこれに倣い,既知個体の識別をクラスレベルの識別とは異なる個体識別能力として扱う(図5.2)。
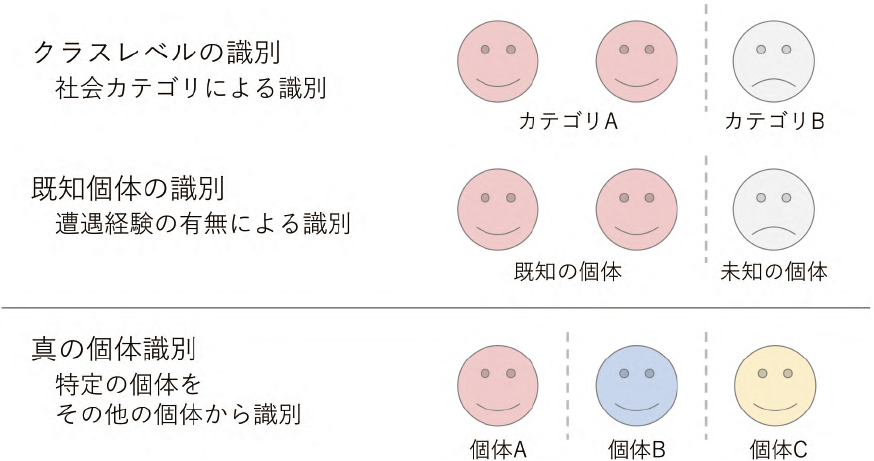
図5.2 各個体識別能力のイメージ。本書では、個体識別を3つに大別する(既知個体の識別をクラスレベルの識別に含める場合もある: 本文)。クラスレベルの識別と既知個体の識別においては、属するカテゴリ ("クラス") が同じである複数個体 (赤の2個体) を区別しない。これら2つの個体識別と比べ、真の個体識別には非常に高度な認知能力が必要となる。
-
-
-
ヤドカリには闘争を介した個体識別能力がありそうだ。しかし,ヤドカリが争うのは貝殻だけではない。オスにとっては成熟したメスも激しい闘争の引き金となる重要な資源である。にもかかわらず,この時点でオス間闘争を介した個体識別能力の研究は存在しなかった。そこでテナガを対象に「オス間闘争における既知個体の識別」の世界初の実証を目指し,博士1年の秋が始まった。
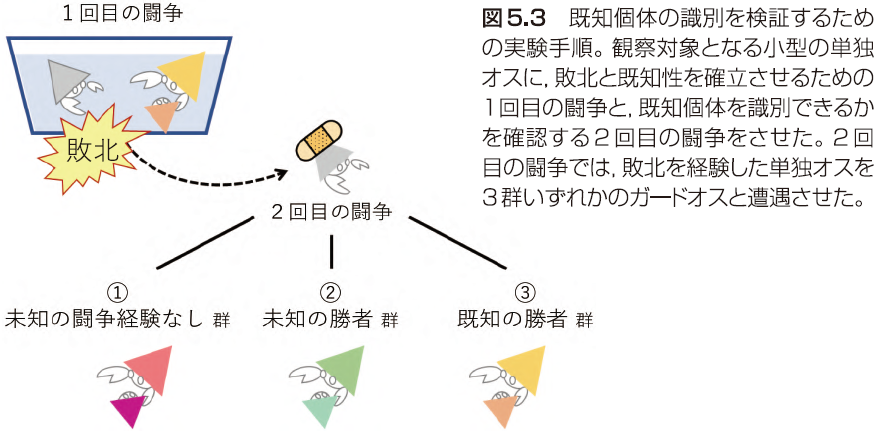
生き物が “闘争相手を個体識別しているか” を検証するには,「相手を覚えるための実験」(1回目の闘争)と,「相手を覚えているか確かめるための実験」(2回目の闘争)が必要である。さらに “既知/未知の個体を識別している” ことを実証するには,「既知の個体に対する行動」と「未知の個体に対する行動」が違うことを示さなければならない。これら一連の実験は少々複雑で,夜のサンプリングを終えた真夜中の頭と体にはいろいろと厳しい。そこで今回は,当時4年生だった後輩3人をお手伝いに任命し,大学で待機してもらった。一方の私は W 先生と夜の海へ。各日の “採れ” 高に当たり外れはあるものの,1度だけ W 先生より1ペア多く採集できたこともあり,交尾前ガードペアの採集スキルは確実に向上していた。誘拐したテナガのペアを抱えて実験室へ戻る。お待たせ後輩たち。これが終われば帰れるぞ。
実験ではまず,3つあるいは2つのペアをランダムに組み合わせた。3つ組では最小の,2つ組では小型のオスからメスを外し,観察対象の単独オスとした。残りのオスはすべてガードオスとする。1回目の闘争では,小型の単独オスに大型オスのペアと 10 分間闘争させて,単独オスに負けてもらった。サイズ上不利な単独オスはふつうガードオスからメスを奪えないので,このオス間闘争は,自分を退けた相手を覚えるための,いわば「やらせ」である(ガードオスからメスを奪えてしまった単独オスは,その後の実験から抜いた)。次に,個体を分けて1時間静置した。最後に,2回目の闘争として,1回目の闘争で負けた単独オスを,①未知の闘争経験なし unfamiliar na¨ıve 群(1回目の闘争に参加していないガードオス。3つ組の余り),②未知の勝者 unfamiliar winner 群(1回目の闘争で別の単独オスを退けたガードオス),③既知の勝者 familiar winner 群(1回目の闘争でその単独オスを退けたガードオス)のいずれかと遭遇させた(図5.3)。
-
-
-
後輩たちと夜な夜な奮闘した結果,テナガのオスが既知個体を識別できる可能性が示唆された(Yasuda et al. 2014a)。1回目の闘争でガードオスに敗北した単独オスは,2回目の闘争相手によって明らかに行動を変えたのである:2回目の闘争で別のガードオスと遭遇した場合,①では1回目の闘争と同じように相手に闘争を仕掛け,②では仕掛けた頻度は若干落ちたものの,粘って闘争を続ける傾向が示された。一方,同じ相手と再闘争させた③では,闘争を仕掛けた単独オスはごくわずかで,仕掛けた後もすぐに撤退した(図5.4)。これらの結果は,本種のオスが,わずか 10 分間のオス間闘争からでも,「1度負けた相手」への既知性を確立し,「そうではない未知の相手」から個体識別することで,勝てる見込みのない闘争を回避できていることを示唆する。
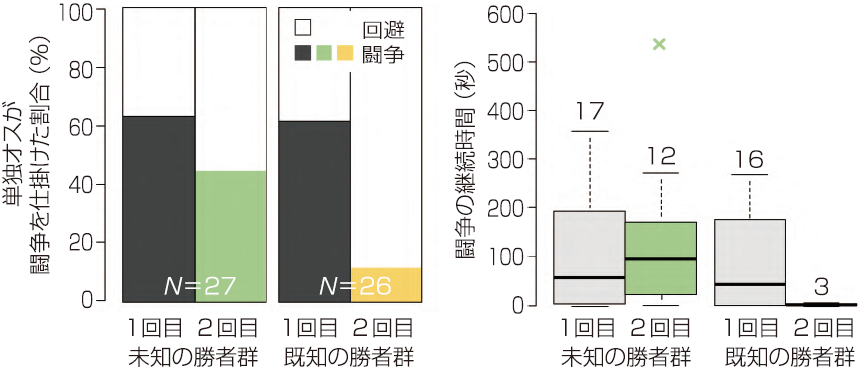
図5.4 テナガホンヤドカリにおける既知個体の識別。未知の勝者群と既知の勝者群を比較した。敗北後の2回目の闘争において、1回目の闘争相手とは異なる未知のガードオスに比べ、1回目の闘争と同じ既知のガードオスと遭遇した単独オスは、相手に闘争を仕掛ける頻度、仕掛けた闘争の継続時間ともに大幅に減少した。箱ひげ図の上の数値は列数を表す。(Yasuda et al. 2014a を基に作成)
-
-
前のコース:卒研論文化までの道のり←|目次|→次のコース: ニュージーランド滞在記