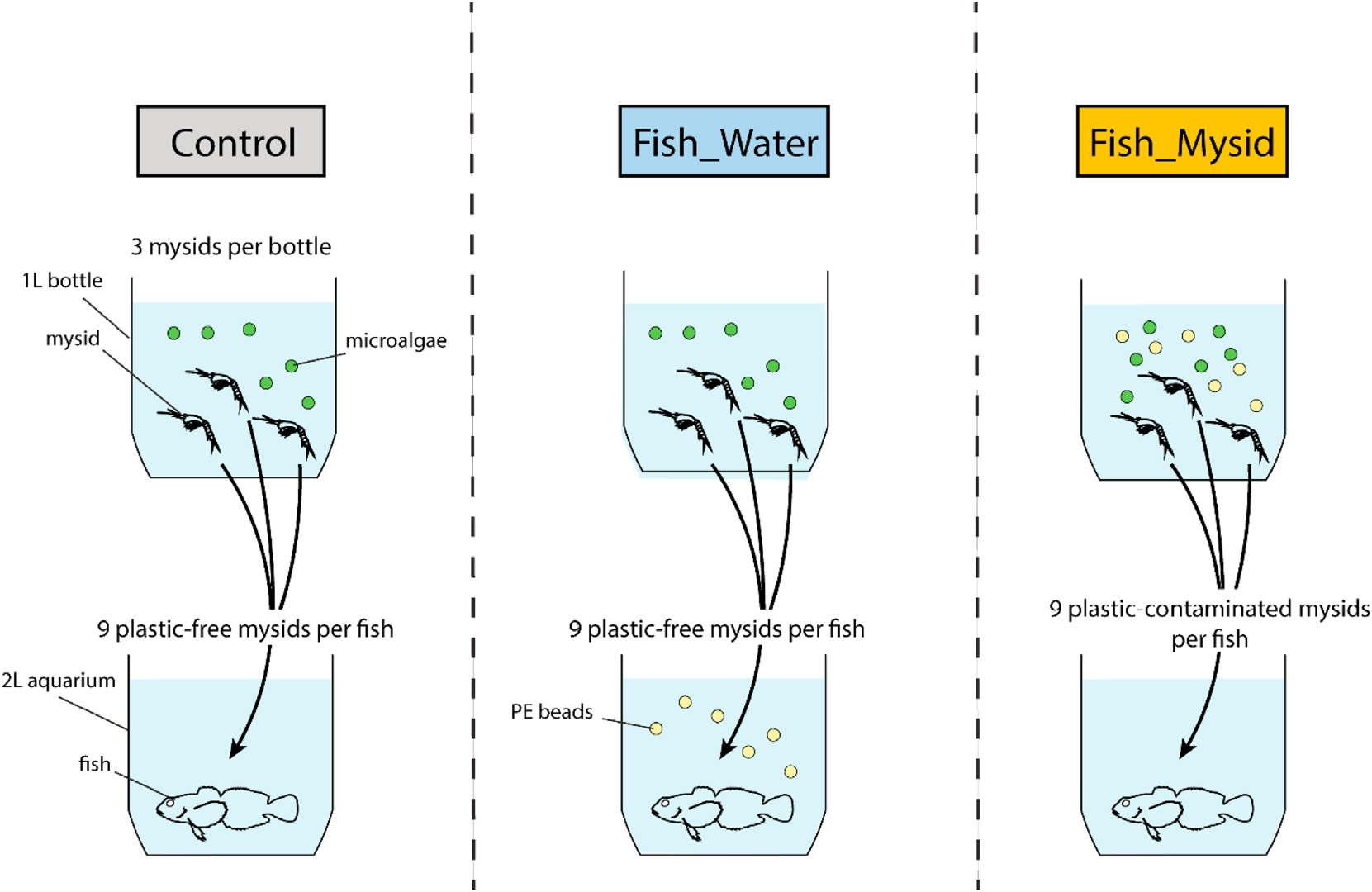
This study evaluated the pathways of microplastic ingestion by fish and biological fragmentation of microplastics by fish and their prey. We found that trophic transfer from prey to predator contributed more than ingestion of microplastics from the water column, and a higher percentage of fragmented plastics was observed in mysids and in the fish that fed on them. To our knowledge, this study is the first to demonstrate that fish ingest more microplastics through their prey than from the water column and that mysids can fragment microplastics.
Most previous studies of microplastics ingestion or toxicity focused on waterborne exposures at the individual level. While there are some empirical studies on trophic transfer of microplastics in aquatic organisms (Elizalde-Velázquez et al., 2020; Nelms et al., 2018; Setälä et al., 2014), none address their contribution relative to ingestion from the water column. In marine fish, microplastic intake via water is considered a major route of exposure (Roch et al., 2020), but our results showed that it is less important than trophic transfer, likely leading to an underestimation of the effects of microplastics in marine food webs. An increase in microplastic ingestion by predator via trophic transfer raises the concerns on their biomagnification. A recent study, however, showed that biomagnification of microplastics are not likely to occur because they can be excreted from an organism and do not accumulate in the body (Walkinshaw et al., 2020). When ingestion rate exceeds the egestion rate, organisms will accumulate microplastics (Au et al., 2017). As trophic transfer contributes to increase the ingestion rate, continuous exposure from the both pathways needs to be considered in future studies to better explore the potential for biomagnification of microplastics.
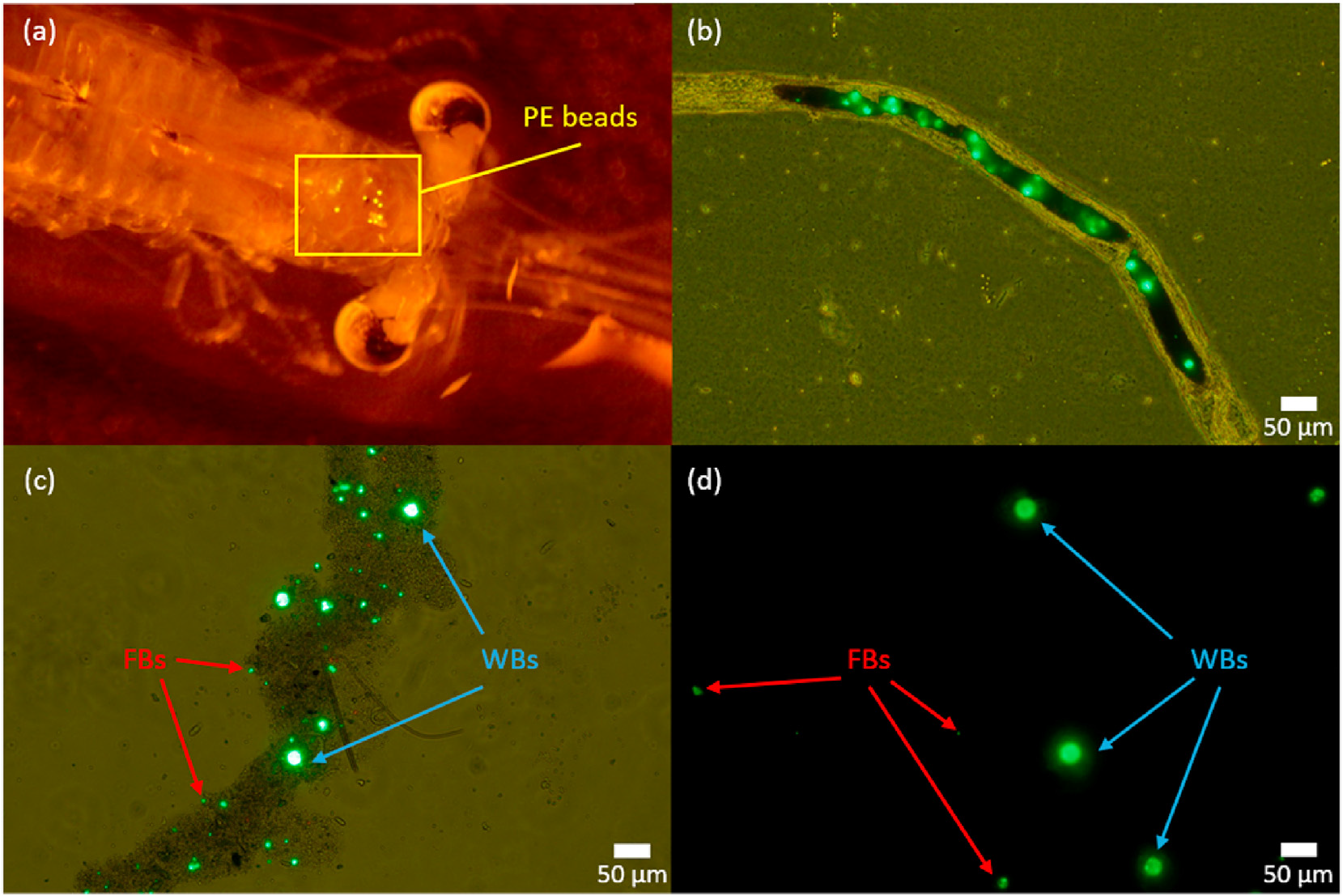
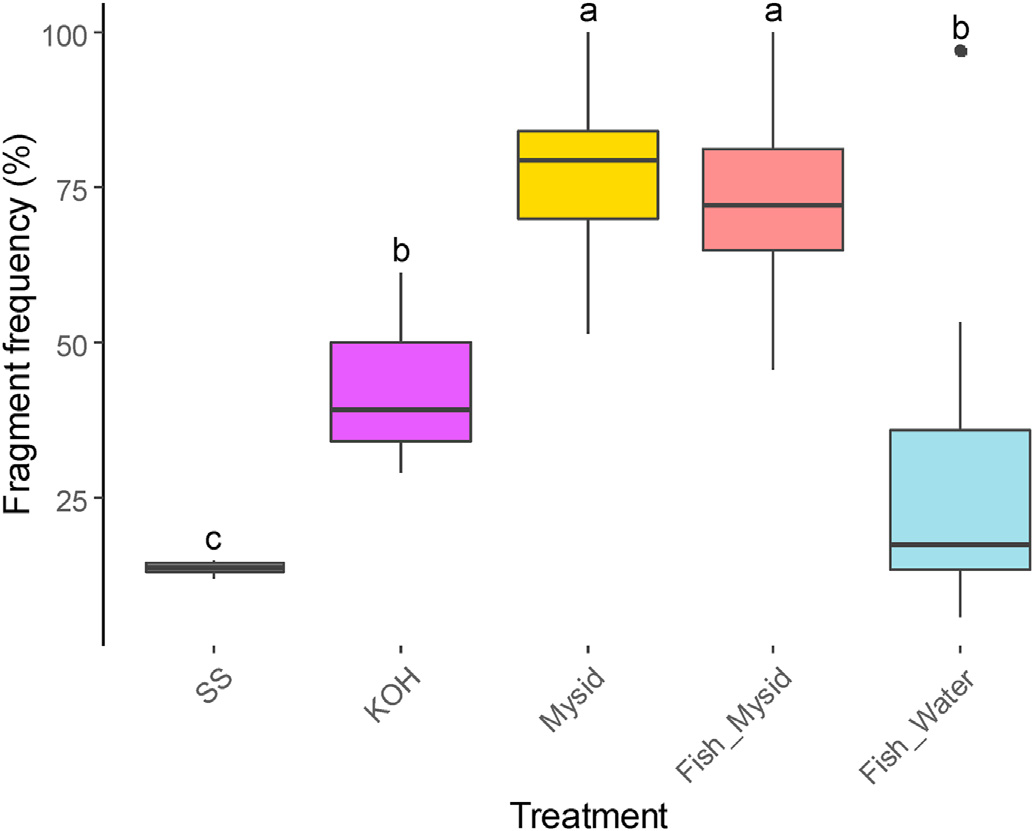
The percentage of smaller plastics recovered from the mysids and fish fed on bead-exposed mysids was significantly higher than in the stock suspension, potassium hydroxide procedure blanks, and fish exposed from the water column. This finding supports our hypothesis that mysids fragment plastics. Dawson et al. (2018) reported that Antarctic krill can fragment microplastics into nanoplastics. An another recent study, by Mateos-Cárdenas et al. (2020), showed that the freshwater amphipod Gammarus duebeni can also fragment microplastics. Many small crustaceans that feed on phytoplankton or detritus, such as krill, amphipods, and mysids, have similar digestive mechanisms, including developed mandibles and chitinous and thick barbed spines in their stomach (Dawson et al., 2018; Friesen et al., 1986; Mateos-Cárdenas et al., 2020). These specific features and mastication in the feeding process explains the capacity for fragmenting plastic particles (Dawson et al., 2018; Mateos-Cárdenas et al., 2020). Although the Antarctic krill used by Dawson and colleagues fragmented over 90% of the plastics ingested, our mysids fragmented only 34% (after subtracting the value in the potassium hydroxide procedure blank), perhaps because of differences in diet. Krill is known to feed mostly on phytoplankton while mysids are omnivorous (Siegfried and Kopache 1980; Nakamura et al., 2020), so krill may have a more developed physical digestive system to grind harder cell structure of phytoplankton. We cannot fully exclude the possibility that the mysids fed selectively on smaller particles, but it is unlikely because mysids preferentially consume phytoplankton exceeding 10 μm in size (Bowers and Grossnickle, 1978; Friesen et al., 1986). Potassium hydroxide fragmented some polyethylene beads, but at a lower frequency than the mysids.
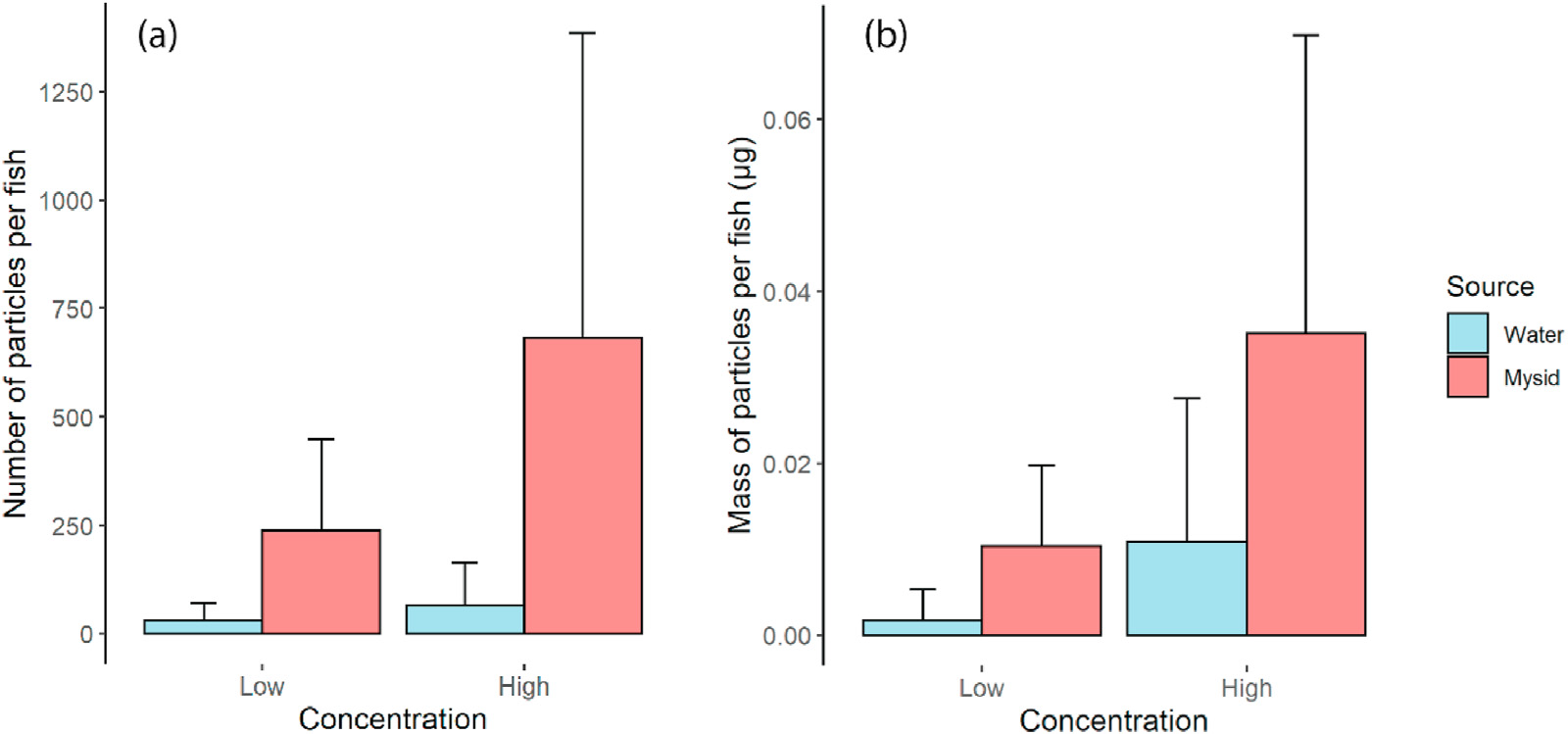
After ingestion by organisms, microplastics can be retained in the digestive tract and cause lacerations, inflammation, and starvation (Carbery et al., 2018). As our study showed, trophic transfer is a significant route of microplastic ingestion and may increase the chance to have such negative impacts on the fish. Particularly, fish exposed to the lower concentration only through the mysids ingested more polyethylene beads than fish exposed to the higher concentration only through the water column. This suggests that trophic transfer of even low concentrations of microplastics have greater impacts on the fish than waterborne ingestion of high concentrations. Yin et al. (2019) demonstrated that waterborne exposure of Korean rockfish (Sebastes schlegelii) to 15-μm polystyrene microplastics altered both energy reserves and behavior. Although larger particles (27-32 μm) were used in our study, the fish fed with bead-exposed mysids contained the more comparable particle size of microplastics (Median: 6.61 μm) to those used in Yin et al. (2019). The concentration used by Yin et al. (2019) was similar to the high-exposure treatment (2000 μg/L, 1.5 × 10⁶ particles) in the current study. As the low-exposure treatment (200 μg/L, 1.5 × 10⁵ particles) is within environmentally relevant concentrations (Goldstein et al., 2012), the actual exposure of predatory fish may be underestimated when it does not take trophic transfer into account. To provide empirical evidence of the importance of trophic transfer in the potential impacts of microplastics on fish, toxicity experiments need to be conducted for more species in future studies.
Our findings also suggest that feeding on small crustaceans such as mysids increases the ingestion of fragmented particles that are small enough to translocate from the digestive tract into other tissues and cause physiological effects (Lu et al., 2016; Von Moos et al., 2012). Although we did not examine the translocation of the fragmented particles into fish tissues, they are theoretically small enough for the translocation to occur. Lu et al. (2016) exposed zebrafish to 5-μmpolystyrene particles and observed accumulation in the liver. The smallest particles observed in our study were 2.43 μm in length, and could potentially translocate to tissues. As we found, feeding on plastic-containing prey increases the number of particles ingested by fish, and the prey can fragment the particles into smaller sizes, so trophic transfer may cause more adverse biological effects in predators.
Plastics present in the oceans contain a variety of toxic chemicals which are either absorbed to the polymer surface from the surrounding water, in particular persistent organic pollutants (POPs; Andrady, 2011) or incorporated during the manufacture process, called plastic additives (Cole et al., 2011). If plastics are ingested by organisms, those chemicals can transfer from the plastics to their tissues and cause adverse health effects (Hermabessiere et al., 2017; Tanaka et al., 2015). Our findings imply that trophic transfer increases microplastic exposure to predators and may accelerate the bioaccumulation of toxic chemicals derived from microplastics. Also, fragmentation of microplastics by small crustaceans could facilitate the release of chemical substances from microplastics due to the migration of the chemicals from the core to the surface of the particle (Wright and Kelly, 2017). Although the biomagnification of microplastics may not occur (Burns and Boxall, 2018; Walkinshaw et al., 2020), plastic-derived chemicals could biomagnify because they can accumulate in organisms’ tissues. More advance research is needed on the effects of trophic transfer and fragmentation of microplastics on the behavior of plastic-derived chemicals.
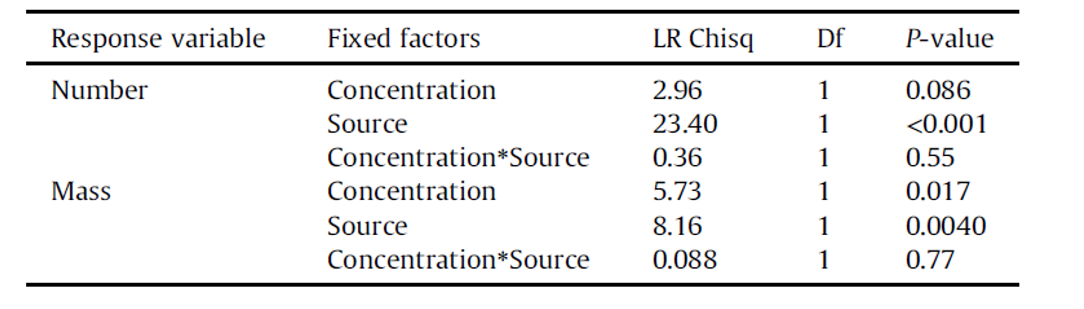
Particle number has been commonly used as a unit of measurement in studies of microplastic ingestion and toxicity, but the lack of standardization precludes direct comparisons between studies. We therefore used particle mass to quantify ingestion and support particle counts. We found that the effect sizes for the difference between ingestion from the water column and ingestion by trophic transfer varied considerably between the number and mass of polyethylene beads. This difference can be attributed to the fragmentation by mysids. As fish ingested fragmented particles through mysids, quantification by number caused an overestimation in the actual quantity of polyethylene beads ingested by the fish, and analysis by mass was more accurate (small: OR < 1.5; medium: 1.5 < OR < 5: large: OR > 5; Chen et al., 2010). Other organisms could also have the capacity to alter the size structure of plastics, then whether showing ingestion values just in the number or mass makes a huge difference in the interpretation of those results. Other organisms may also fragment plastic microspheres, and we therefore strongly recommend that results be analyzed by mass and not by particle number for more accurate quantification.