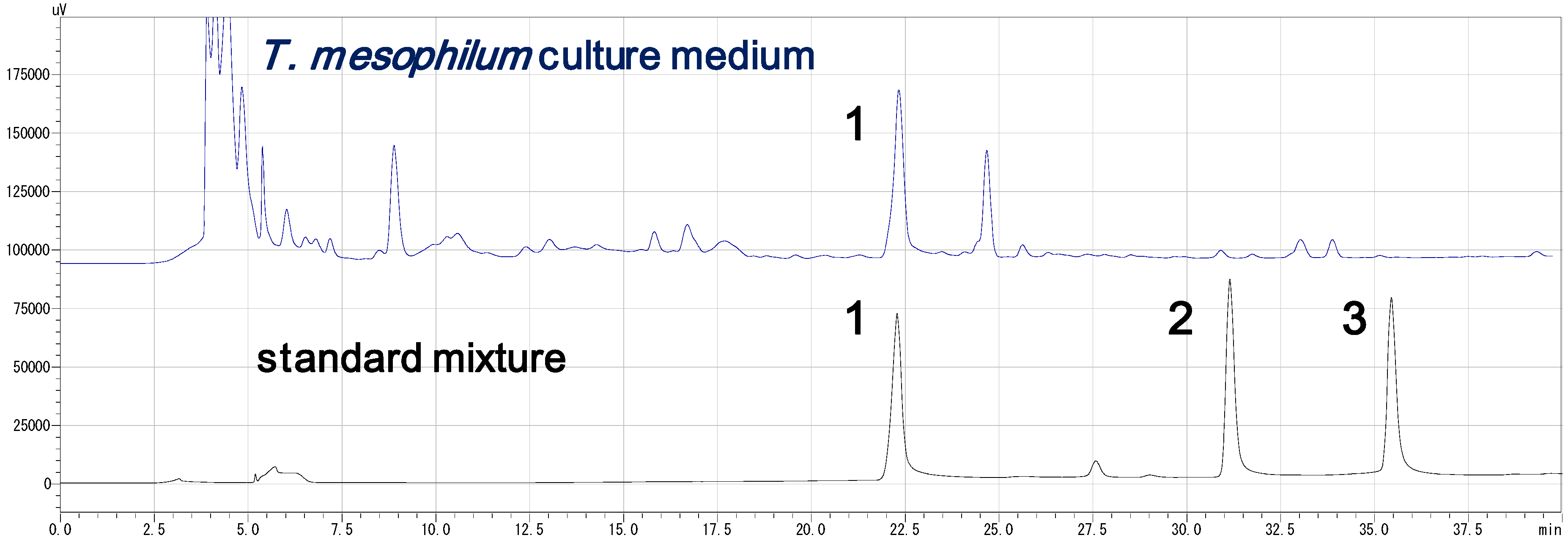
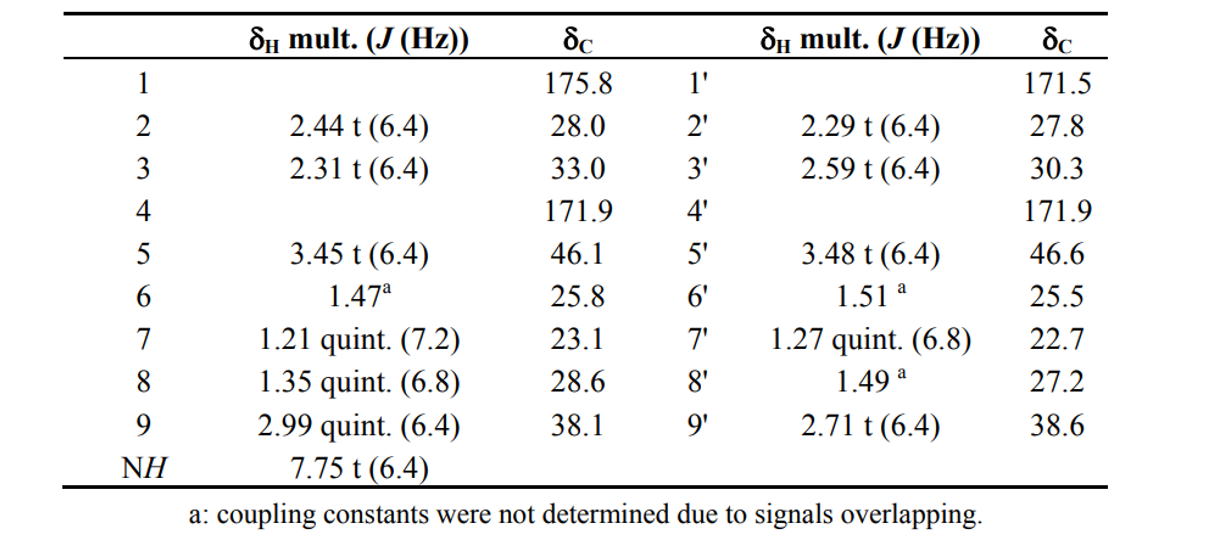
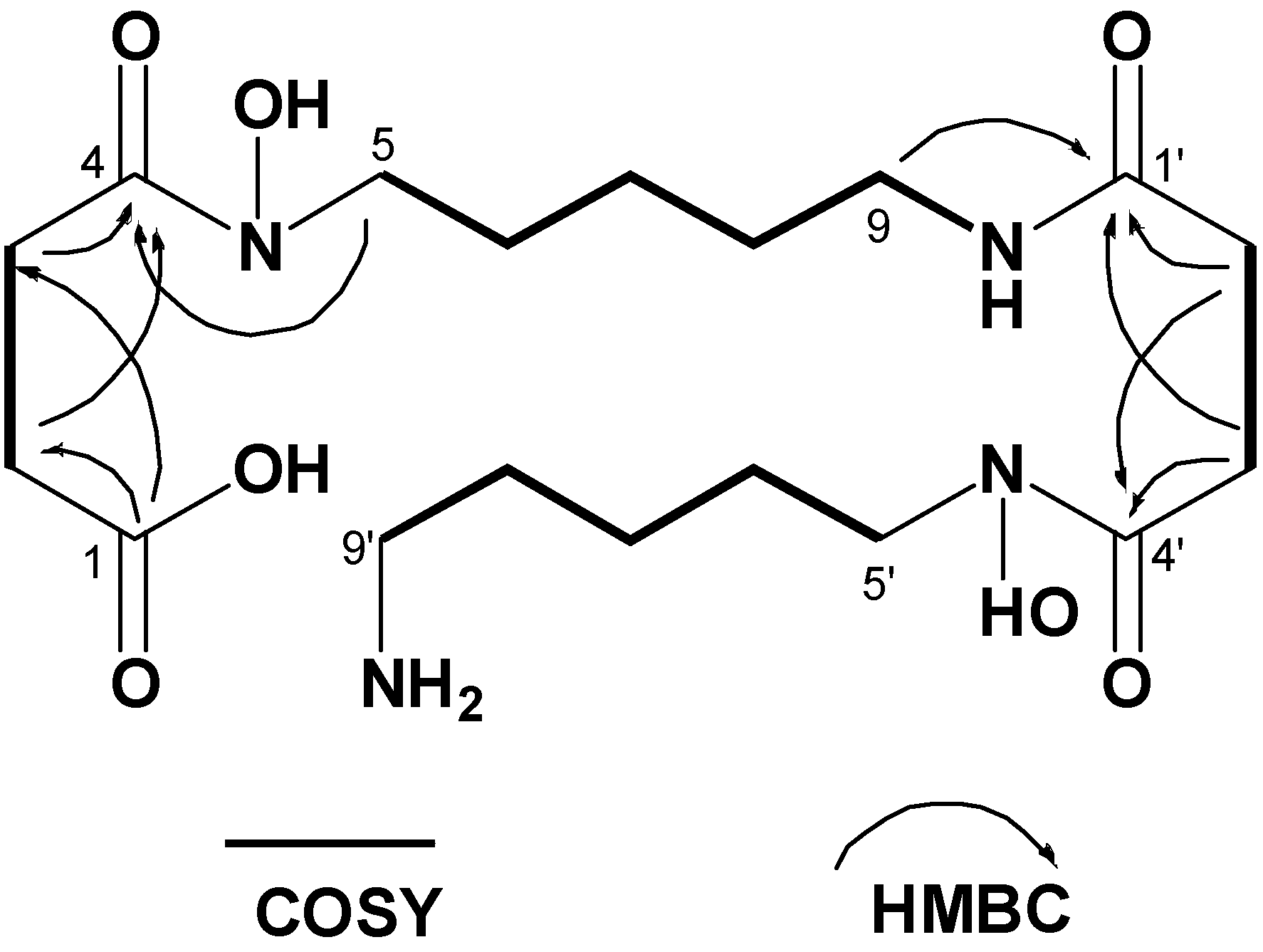
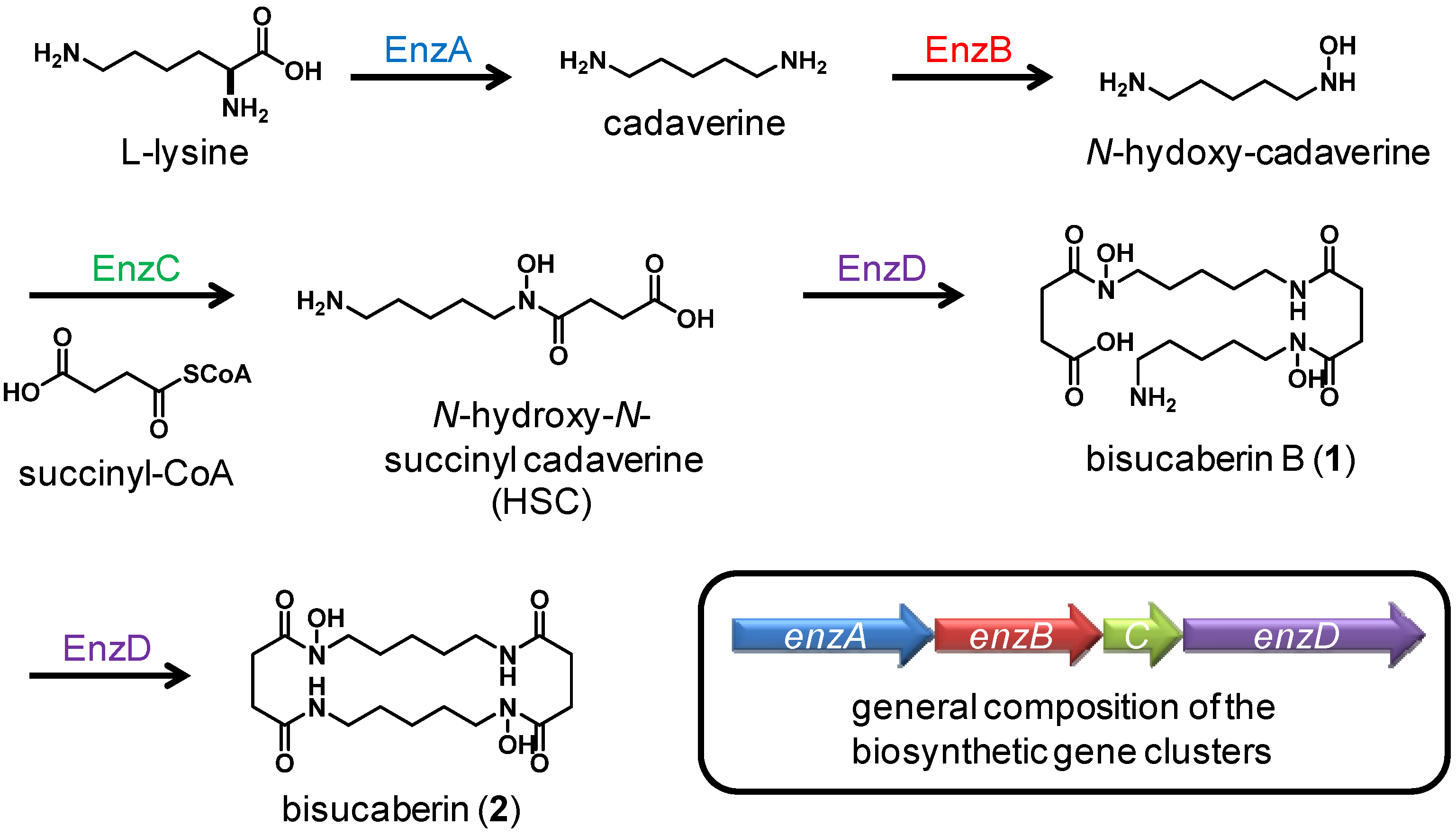
Bisucaberin B (1)は、以前、desferrioxamine Bのバクテリア分解によるマイナー代謝物として報告されました(4) 。今回、我々はbisucaberin B(1)が唯一のsiderophoreとしてデノボに生合成されることを初めて明らかにしました。これまでに、Streptomyces属、Pseudomonas属、Erwinia属、Vibrio属の細菌から、3量体のアナログであるdesferrioxamine G1(5)が報告されています。しかし、ほとんどの場合、化合物5は大環状化合物3と共生成されており、化合物3の生合成中間体であると考えられています。本研究で培養液中に環状生成物の痕跡が認められなかったことは、化合物1が環状生成物の加水分解物ではなく、直鎖状の最終生成物として形成されることを強く示唆しています(図2)。
図5に、HSC系siderophoreの一般的な生合成スキームと生合成遺伝子クラスターの構成を示します。最初の3つの酵素EnzA-Cはモノマー生成を触媒し、最後の酵素EnzDはオリゴマー化と大環状化反応の両方を触媒します。T. mesophilumが直鎖状の二量体のみを生成し、環状生成物を生成しないことから、この株の酵素Dはモノマー間のアミド結合の生成を触媒し、大環状化を触媒しないことが示唆され、これが事実ならば、これまでに報告されていない現象です。酵素Dを他の既知の大環状化酵素と比較することで、大環状化反応の分子機構が明らかになると期待されます。オリゴマー化-大環状化機構を深く理解することで、所望のサイズと構造を持つHSCベースのsiderophoreの人工的な生産が可能になると期待されます。
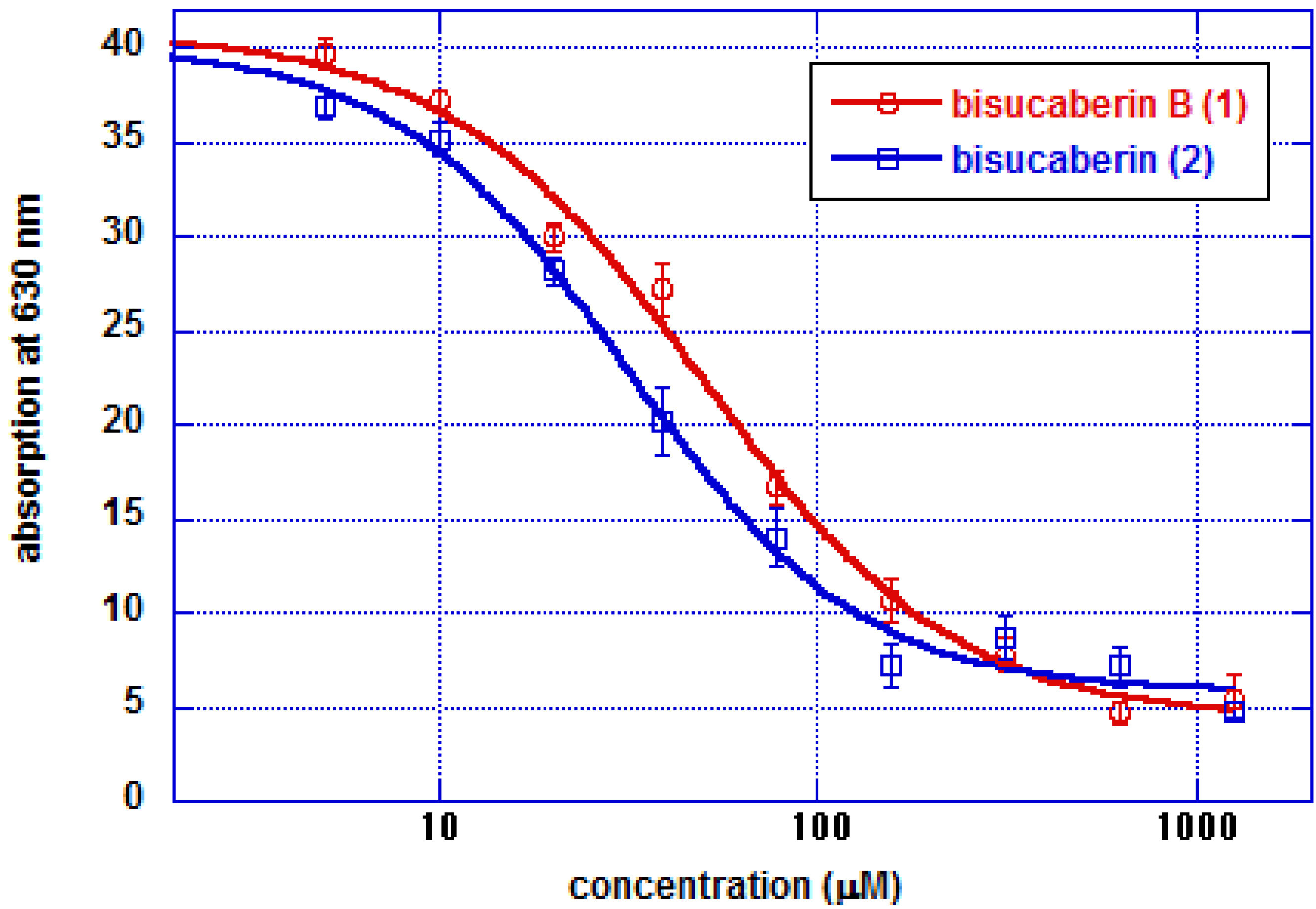
直鎖状二量体の鉄イオンキレート活性は、大環状二量体のそれと同等でした。我々は、非環状キレーターの生産は、他のバクテリアによるsiderophoreの改変行為を防ぐための対策であると仮定しています。化合物1と2のsiderophore-鉄イオン複合体のコンフォメーションは異なると予想されるため、鉄複合体化した化合物1は、外来siderophoreを改変行為する近隣の細菌が発現する化合物2の受容体に認識されない可能性があります。このように、直鎖型および環状HSC系siderophoreに対するsiderophore受容体の選択性を比較することは、化学生態学の観点から興味深いと思われます。化合物1の生合成酵素および推定受容体をコードする遺伝子のクローニングが進行中です。